More Than Just a FAD(5): Unsaturated Fatty Acids in Chloroplasts Elicit Protective Autoimmunity
Arabidopsis (Arabidopsis thaliana) chloroplast-division mutants that have abnormally large chloroplasts have been around for quite some time, not only because they can be identified relatively easily through screening (Pyke & Leech 1991) and because their spectacular morphology sparks intrigue, but because they have proven useful to this day for studying several important biological phenomena related not just to chloroplast division, but to control of cell size, chloroplast movement and light avoidance, and photosynthesis (Dutta et al. 2017). “Giant chloroplast” (GC) mutants, including accumulation and replication of chloroplasts 6 (arc6) and crumpled leaf (crl), also exhibit light-induced localized cell death (LCD). However, the mechanism controlling this phenotype had remained elusive, until Li et al. (2020) recently discovered the involvement of chloroplast-localized FATTY ACID DESATURASE5 (FAD5).
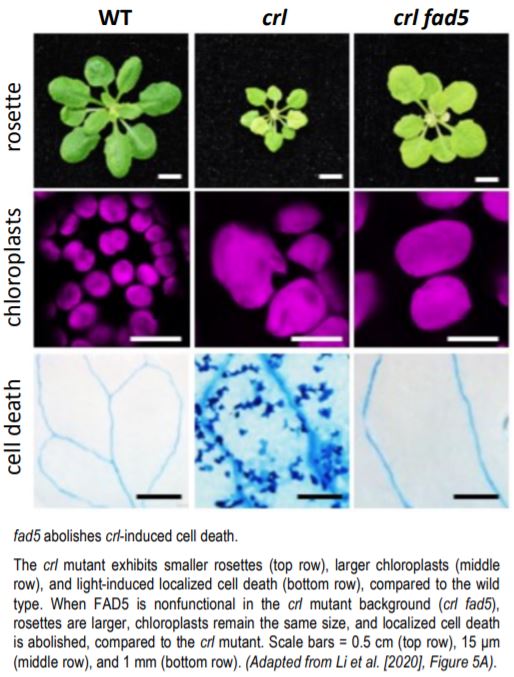 The fad5 mutation was identified in a forward-genetics suppressor screen of the crl mutant. fad5 completely rescued the LCD and partially rescued the small rosette and crumply leaves phenotypes of the crl mutant; however, the chloroplasts remained large (see figure). fad5 also dampened the crl-induced upregulation of stress- and detoxification-induced nuclear genes. Likewise, when fad5 was introduced into the backgrounds of other plants with genetically determined GCs, such as arc6, a similar reduction in both LCD and the expression of stress-related genes resulted, while the large chloroplast size remained intact. Thus, it seems that there is a common signaling pathway to LCD that is initiated by GCs, and that FAD5 is involved—but how?
The fad5 mutation was identified in a forward-genetics suppressor screen of the crl mutant. fad5 completely rescued the LCD and partially rescued the small rosette and crumply leaves phenotypes of the crl mutant; however, the chloroplasts remained large (see figure). fad5 also dampened the crl-induced upregulation of stress- and detoxification-induced nuclear genes. Likewise, when fad5 was introduced into the backgrounds of other plants with genetically determined GCs, such as arc6, a similar reduction in both LCD and the expression of stress-related genes resulted, while the large chloroplast size remained intact. Thus, it seems that there is a common signaling pathway to LCD that is initiated by GCs, and that FAD5 is involved—but how?
The crl mutant makes a lot of singlet oxygen (1O2; a type of reactive oxygen species [ROS]), its source presumably the GCs caused by impaired chloroplast division. Concomitantly, the crl mutant constitutively expresses 1O2-responsive genes. Although salicylic acid (SA)- and jasmonic acid (JA)-responsive genes in particular were upregulated in the crl mutant, and ROS can signal through these stress hormones, neither SA nor JA seem to be involved in eliciting crl-specific LCD. When SA or JA signaling was genetically depleted in the crl mutant, the crl-induced LCD phenotype was not rescued. Additionally, no effect on LCD or other phenotypes in the crl mutant background was observed when 1O2 signaling was impaired via loss of EXECUTER proteins, which sense or modulate signaling of 1O2.
However, biological molecules, including lipids, react with 1O2. Polyunsaturated fatty acids (PUFAs), but not saturated fatty acids, are particularly susceptible to oxidation, leading to lipid peroxidation and/or reactive electrophile species (RES). Notably, the crl mutant exhibited an upregulation of RES-responsive genes. Since FAD5 specifically converts palmitic acid (a 16:0 saturated fatty acid) to palmitoleic acid (a 16:1 unsaturated fatty acid) in the chloroplast, it is able to create a population of fatty acids with which 1O2 can react. Thus, when the crl and other GC mutants lack functional FAD5, these resulting RES and/or lipid peroxides decrease, leading to reduction of LCD. As expected, the crl fad5 mutant had significantly higher levels of C16:0 and lower levels of C16:1 compared to the crl mutant. Although the lipid profile of the crl fad5 mutant was clearly altered, the LCD abolishment was not simply due to perturbed lipid homeostasis, as altering the profile in a different way by generating a crl fad7 fad8 mutant had no effect on LCD, suggesting that FAD5 is specifically responsible for LCD in the crl mutant. It turns out, therefore, that particular PUFAs that get oxidized by 1O2 as a result of ROS production in chloroplasts can serve as a beacon that induces autoimmune responses.
Li et al. (2020) highlight, once again, the utility of GC mutants in studying biological phenomena—this time, autoimmunity. Intriguingly, there are other drawbacks to having large chloroplasts. It was previously found that plants with larger-than-normal chloroplasts accumulate more photosystem II photodamage, independent of their chloroplast movement impairment (Dutta et al. 2017). However, the fad5 mutation does not seem to have any alleviating effect on photosynthetic activity in the crl mutant. There is clearly more to learn. But because of the negative consequences that GCs have to overall plant size and photosynthetic capacity, GC mutants are certainly here to stay in plant biology research!
Anne C. Rea
Michigan State University
MSU-DOE Plant Research Laboratory
ORCID: 0000-0002-2996-5709
REFERENCES
Dutta, S., Cruz, J.A., Imran, S.M., Chen, J., Kramer, D.M., and Osteryoung, K.W. (2017). Variations in chloroplast movement and chlorophyll fluorescence among chloroplast division mutants under light stress. J. Exp. Bot. 68: 3541–3555.
Pyke, K.A. and Leech, R.M. (1991). Rapid image analysis screening procedure for identifying chloroplast number mutants in mesophyll cells of Arabidopsis thaliana (L.) Heynh. Plant Physiol. 96: 1193–1195.