A new order through disorder: intrinsically disordered proteins reshape the cytoskeleton under drought stress
Many proteins have a clearly defined structure-function relationship – specific three-dimensional folds can be linked to specific protein functions. We do however know that many proteins – referred to as intrinsically disordered proteins (IDPs) – lack rigid three-dimensional organisation. In recent years, IDPs have been recognised as key components of many signalling networks. In plants, they are linked particularly to stress responses, during which they perform important functions to elicit appropriate physiological and developmental changes that are needed for environmental acclimation and survival (Covarrubias et al., 2017). Intrinsic disorder allows these proteins to interact with multiple partners, but also to form highly complementary binding interfaces that ensure high binding specificity. Moreover, while many proteins tend to aggregate under adverse conditions such as heat or water shortage, their flexible structure renders IDPs generally less prone to aggregation. Taken together, these features make them excellent candidates for signalling hubs under stress conditions (Sun et al., 2013; Liu and Huang, 2014). While many IDPs are associated with plant stress responses, our understanding of the underlying signalling mechanisms is still fairly limited. In this issue of Plant Physiology, Hsiao et al. (2020) reveal how a group of rice (Oryza sativa) IDPs contributes to changes in root architecture under drought stress.
 Upon perception of water shortage, plants trigger an array of responses to ensure survival, ranging from short-term physiological acclimation such as stomatal closure to long-term alterations in their development. Growth usually slows down when water is scarce, and although root growth is usually less inhibited than shoot growth, prolonged water shortage does eventually cause progressive inhibition of root elongation (Sharp et al., 2004). Both root growth maintenance and growth arrest are regulated by the phytohormone abscisic acid (ABA) (Sharp et al., 2004; Sun et al., 2018). Accumulation of ABA is a widespread response to drought and in turn triggers expression of many stress-associated genes. In rice, expression of the REPETITIVE PROLINE-RICH PROTEIN (OsRePRP) gene family is strongly upregulated in the root elongation zone upon water stress and ABA treatment, and the respective proteins are required to restrict root elongation growth under severe drought stress (Tseng et al., 2013).
Upon perception of water shortage, plants trigger an array of responses to ensure survival, ranging from short-term physiological acclimation such as stomatal closure to long-term alterations in their development. Growth usually slows down when water is scarce, and although root growth is usually less inhibited than shoot growth, prolonged water shortage does eventually cause progressive inhibition of root elongation (Sharp et al., 2004). Both root growth maintenance and growth arrest are regulated by the phytohormone abscisic acid (ABA) (Sharp et al., 2004; Sun et al., 2018). Accumulation of ABA is a widespread response to drought and in turn triggers expression of many stress-associated genes. In rice, expression of the REPETITIVE PROLINE-RICH PROTEIN (OsRePRP) gene family is strongly upregulated in the root elongation zone upon water stress and ABA treatment, and the respective proteins are required to restrict root elongation growth under severe drought stress (Tseng et al., 2013).
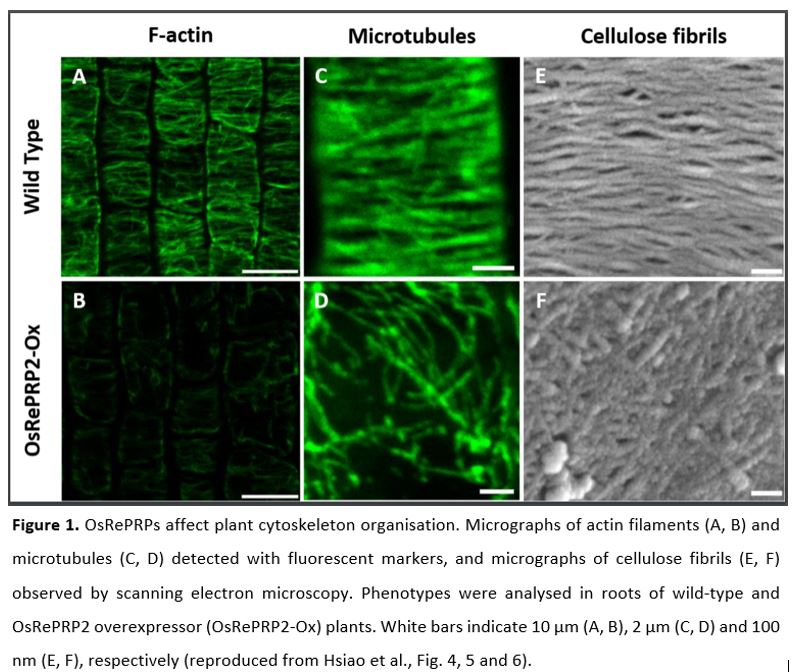
OsRePRPs are predicted to be intrinsically disordered, with a proline content of up to 40% (Tseng et al., 2013), and these proline-rich domains bear resemblance to actin-binding proteins in animals. This observation prompted Hsiao et al. to investigate whether OsRePRPs interact with actin as well as tubulin, the other main component of the plant cytoskeleton. They indeed observed interaction of OsRePRP1 and OsREPRP2 with both actin and tubulin in vitro and in vivo and found OsRePRPs to co-localise with cytoskeleton markers in planta. Taken together, these observations imply a functional link between OsRePRPs and the plant cytoskeleton. While the cytoskeleton takes on manifold functions, in plant cells it is particularly important for cell-wall structure and composition: actin filaments provide “roads” for vesicle trafficking to and from the plasma membrane while microtubules guide cellulose synthase complexes and thereby determine the orientation of cellulose fibrils within the cell wall. Do OsRePRPs affect these processes? To answer this question, the authors analysed multiple molecular and cellular phenotypes in OsRePRP2 overexpressor and OsRePRP1/2 RNAi knock-down lines. Their analyses revealed that OsRePRPs reduce the abundance of actin filaments (Fig. 1A, B), causing reduced transport of non-cellulosic polysaccharides to the plasma membrane. In parallel, OsRePRPs trigger reorientation of microtubules, which translates into a disorganised network of cellulose microfibrils in the cell wall (Fig. 1C-F). Together, these effects hamper cell expansion and thereby restrict elongation growth in roots.
Notably, Hsiao et al. find that severe water shortage and OsRePRP overexpression make roots not only shorter, but also heavier – OsRePRPs boost starch accumulation, which is potentially linked to increased sucrose synthase activity, and thereby cause roots to “gain weight”. Therefore, it seems that under severe drought stress, rice roots switch from source to sink mode – they arrest growth and become a dormant storage organ. Entering a quiescent state is also observed under what could be considered the opposite condition, namely prolonged submergence. The molecular response to flooding is quite distinct from the drought response and involves accumulation of the plant hormone ethylene (Loreti et al., 2016). While the signalling cascades leading to it differ considerably under different circumstances, quiescence may represent a general strategy of certain rice cultivars to overcome unfavourable environmental conditions and ensure plant survival.
In conclusion, the study by Hsiao et al. uncovers a molecular role for IDPs of the OsRePRP family, which can reorganise the highly ordered structure of a cell’s cytoskeleton to alter root growth and physiology to promote stress tolerance. Especially the fact that they can interact with both major types of plant cytoskeleton filaments is quite remarkable and certainly warrants further investigation. It remains to be seen how exactly the OsRePRPs establish these interactions and thereby trigger cytoskeletal reorganisation; a combination of cell biological, biophysical and biochemical approaches will be required to fully understand the underlying mode of action of these proteins.
Martin Balcerowicz
Assistant Features Editor
Sainsbury Laboratory, University of Cambridge, Cambridge CB2 1LR, United Kingdom
Literature Cited
Covarrubias AA, Cuevas-Velazquez CL, Romero-Pérez PS, Rendón-Luna DF, Chater CCC (2017) Structural disorder in plant proteins : where plasticity meets sessility. Cellular and Molecular Life Sciences 74: 3119–3147
Hsiao A-S, Wang K, H T-HD (2020) Intrinsically Disordered Protein Interacts with Cytoskeleton for Adaptive Root Growth under Stress. Plant Physiology https://doi.org/10.1104/pp.19.01372
Liu Z, Huang Y (2014) Advantages of proteins being disordered. Protein Sci 23: 539–550
Loreti E, van Veen H, Perata P (2016) Plant responses to flooding stress. Current Opinion in Plant Biology 33: 64–71
Sharp RE, Poroyko V, Hejlek LG, Spollen WG, Springer GK, Bohnert HJ, Nguyen HT (2004) Root growth maintenance during water deficits: physiology to functional genomics. J Exp Bot 55: 2343–2351
Sun X, Rikkerink EHA, Jones WT, Uversky VN (2013) Multifarious Roles of Intrinsic Disorder in Proteins Illustrate Its Broad Impact on Plant Biology. The Plant Cell 25: 38–55
Sun LR, Wang YB, He SB, Hao FS (2018) Mechanisms for Abscisic Acid Inhibition of Primary Root Growth. Plant Signaling & Behavior 13: e1500069
Tseng I-C, Hong C-Y, Yu S-M, Ho T-HD (2013) Abscisic Acid- and Stress-Induced Highly Proline-Rich Glycoproteins Regulate Root Growth in Rice. Plant Physiology 163: 118–134



")



