An essential role of MADS-box proteins during wheat spike development (Plant Cell)
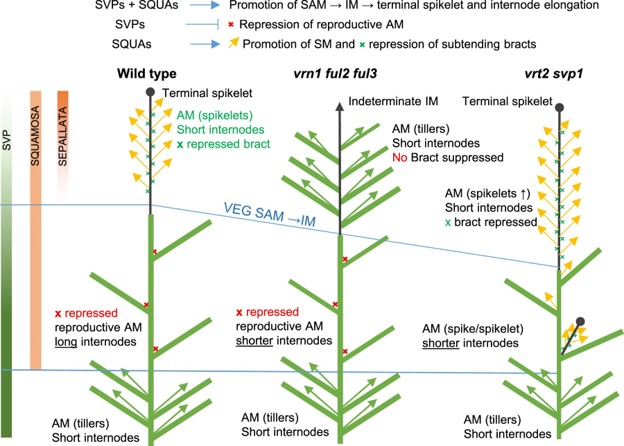 Wheat yields depend on the morphology of the inflorescence known as the spike, which harbors multiple lateral spikelets. To control and manipulate wheat productivity, it is necessary to understand the molecular and genetic mechanisms behind the development of inflorescence architecture. Wheat spike development from the inflorescence meristem (IM) is determined when the terminal spikelet (TS) is formed. The transition from IM to TS largely influences the number of spikelets and eventually the number of grains per spike. Two genes of the SQUAMOSA clade of the MADS-box TF family, i.e., VERNALIZATION1 (VRN1) and FRUITFULL2 (FUL2), are required for this transition, and consequently, spikelets development. In the vrn1ful2 double mutant, the development of TS is abolished, and the axillary meristem generates tiller-like vegetative structures instead of generating spikelets. However, the single mutant (vrn1 or ful2) spikes develop normally with more spikelets due to delayed transition to TS. To explore the regulatory network involved in the spike development, Li et al. (2021) compared the transcriptomes of developing spikes of vrn1 and vrn1ful2 mutants. This led to the identification of three MADS-box genes of SVP clade, i.e., SHORT VEGETATIVE1 (SVP1), SVP2/VEGETATIVE TO REPRODUCTIVE TRANSITION2 (VRT2), and SVP3. Further analysis using genetic interactions among SQUAMOSA and SVP genes revealed their synergistic effects on IM to TS transition but antagonistic effects on the development of spikelets from axillary meristems. SQUAMOSA genes control spikelet development from axillary meristem by repressing SVP genes. Altogether, this SQUAMOSA-SVP cascade is essential for TS development, and the downregulation of SVP genes is required for normal spikelet development. (Summary by Subodh Verma @subodhshanky) Plant Cell 10.1093/plcell/koab243.
Wheat yields depend on the morphology of the inflorescence known as the spike, which harbors multiple lateral spikelets. To control and manipulate wheat productivity, it is necessary to understand the molecular and genetic mechanisms behind the development of inflorescence architecture. Wheat spike development from the inflorescence meristem (IM) is determined when the terminal spikelet (TS) is formed. The transition from IM to TS largely influences the number of spikelets and eventually the number of grains per spike. Two genes of the SQUAMOSA clade of the MADS-box TF family, i.e., VERNALIZATION1 (VRN1) and FRUITFULL2 (FUL2), are required for this transition, and consequently, spikelets development. In the vrn1ful2 double mutant, the development of TS is abolished, and the axillary meristem generates tiller-like vegetative structures instead of generating spikelets. However, the single mutant (vrn1 or ful2) spikes develop normally with more spikelets due to delayed transition to TS. To explore the regulatory network involved in the spike development, Li et al. (2021) compared the transcriptomes of developing spikes of vrn1 and vrn1ful2 mutants. This led to the identification of three MADS-box genes of SVP clade, i.e., SHORT VEGETATIVE1 (SVP1), SVP2/VEGETATIVE TO REPRODUCTIVE TRANSITION2 (VRT2), and SVP3. Further analysis using genetic interactions among SQUAMOSA and SVP genes revealed their synergistic effects on IM to TS transition but antagonistic effects on the development of spikelets from axillary meristems. SQUAMOSA genes control spikelet development from axillary meristem by repressing SVP genes. Altogether, this SQUAMOSA-SVP cascade is essential for TS development, and the downregulation of SVP genes is required for normal spikelet development. (Summary by Subodh Verma @subodhshanky) Plant Cell 10.1093/plcell/koab243.