The Power of Plasticity in Polyploid Persimmon
IN BRIEF by Jennifer Lockhart [email protected]
Most plants are hermaphrodites, producing perfect flowers with both male and female functions. In roughly 6% of plants, however, male (usually XY) plants produce only male flowers and female (XX) plants produce only female flowers. These dioecious plants cannot self-pollinate, ensuring genetic diversity and facilitating the breeding process. In the dioecious diploid persimmon (Diospyros lotus), sex is determined by the autosomal gene MeGI (Japanese for female tree) and the Y-encoded pseudogene OGI (male tree) (Akagi et al., 2014). MeGI encodes a homeodomain transcription factor that regulates flower development and anther fertility in a dosage-dependent manner, acting as a feminizing agent. In females, MeGI builds to high levels, repressing pollen formation. In males, OGI produces a small RNA (smRNA) that targets and represses MeGI, thereby preventing the accumulation of MeGI.
The situation is more complex in the autohexaploid persimmon species D. kaki. These trees are either fully female or monoecious (male and female flowers on separate branches). Monoecious trees are genetically male, prompting Akagi et al. (2016) to wonder how plants with Y chromosomes produce female flowers. Their analysis uncovered an intriguing epigenetic regulatory mechanism not found in D. lotus.
The authors began by comparing three types of flowers: female flowers from female trees, female flowers from monoecious trees, and male flowers from monoecious trees. Like D. lotus flowers, male D. kaki flowers harbor MeGI targeted by smRNA, with all six copies of this gene equally targeted. As expected, OGI expression was completely undetectable in female flowers from both female and monoecious D. kaki trees. Surprisingly, OGI expression was also undetectable in male flowers from monoecious trees. It turns out that the OGI promoter in D. kaki is interrupted by a 268 bp insertion resembling a SINE (short interspersed nuclear element)-like retrotransposon element, which appears to be unique to monoecious D. kaki cultivars. This element exhibits heavy smRNA accumulation, along with strong cytosine methylation, in both male and female buds and flowers from genetically male plants, suggesting it represses OGI expression in D. kaki.
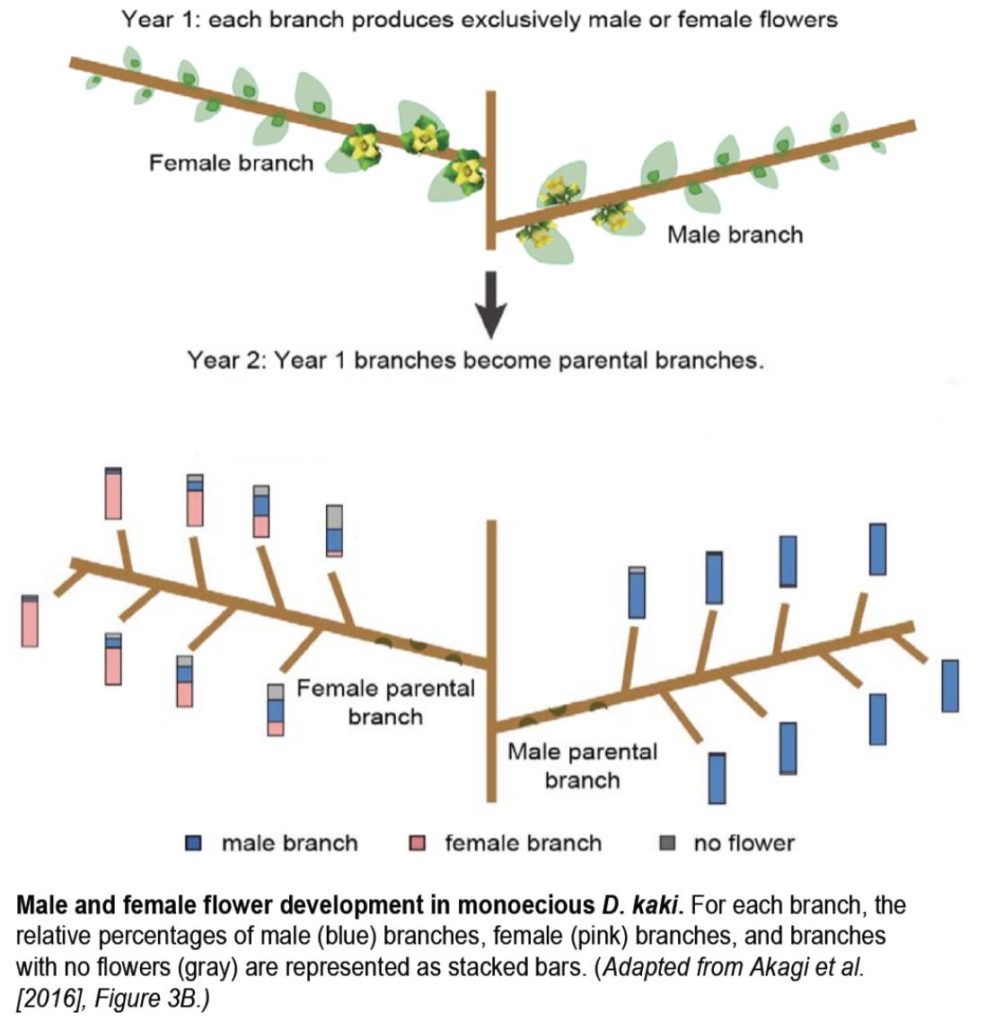 The position of a D. kaki bud on a branch affects the sex of the resulting flowers the following year (see figure). In June, each branch bears only male or female flowers and developing buds of unknown sex. The following year, the buds on these male or female parental branches develop into new, secondary branches, each bearing only male or female flowers. Apical buds on female branches almost always give rise to female secondary branches, whereas medial buds on the same branches have a nearly equal chance of forming male or female branches. By contrast, buds from male parental branches almost always develop into male branches, regardless of their position on the parental branch. This pattern suggests that an epigenetic mechanism is at play. Indeed, the methylation pattern across MeGI in D. kaki strongly mirrors this trend, with the highest methylation levels found in mature male flowers and buds and the lowest in their female counterparts. Therefore, gene silencing by methylation appears to regulate sex determination in D. kaki. This differential methylation pattern is much stronger in D. kaki than in D. lotus, which mainly uses OGI to silence this feminizing gene, suggesting that methylation of MeGI represents an alternative mechanism for sex determination in this autohexaploid persimmon species.
The position of a D. kaki bud on a branch affects the sex of the resulting flowers the following year (see figure). In June, each branch bears only male or female flowers and developing buds of unknown sex. The following year, the buds on these male or female parental branches develop into new, secondary branches, each bearing only male or female flowers. Apical buds on female branches almost always give rise to female secondary branches, whereas medial buds on the same branches have a nearly equal chance of forming male or female branches. By contrast, buds from male parental branches almost always develop into male branches, regardless of their position on the parental branch. This pattern suggests that an epigenetic mechanism is at play. Indeed, the methylation pattern across MeGI in D. kaki strongly mirrors this trend, with the highest methylation levels found in mature male flowers and buds and the lowest in their female counterparts. Therefore, gene silencing by methylation appears to regulate sex determination in D. kaki. This differential methylation pattern is much stronger in D. kaki than in D. lotus, which mainly uses OGI to silence this feminizing gene, suggesting that methylation of MeGI represents an alternative mechanism for sex determination in this autohexaploid persimmon species.
Treatment with the non-specific methylation inhibitor zebularine helped confirm this model: zebularine treatment tended to promote pistil development and inhibit anther development in male flowers of male-based monoecious D. kaki cultivars, but it had no effect on female flowers. Methylation
marks could also be removed in nature, inducing a switch from male to female flower production. Such plasticity would allow plants to optimize female flower (and fruit) production based on environmental conditions. This elegant mechanism has possible implications for fruit production in D. kaki. More generally, this work sheds light on the influence of methylation on gene function and phenotypic variation in polyploids.
REFERENCES
Akagi, T., Henry, I.M, Kawai, T., Comai, L., and Tao, R. (2016). Epigenetic regulation of the sex determination gene MeGI in polyploid persimmon. Plant Cell 28: doi:10.1105/tpc.16.00532.
Akagi, T., Henry, I.M., Tao, R., and Comai, L. (2014). A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 346: 646–650.







Leave a Reply
Want to join the discussion?Feel free to contribute!