Update: Cation/H+ transporters affect membrane trafficking
 By
By
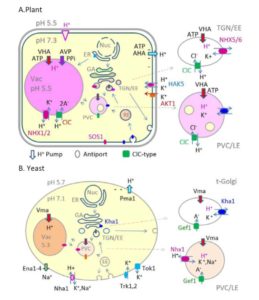
Plants remodel their cells through the dynamic endomembrane system. Intracellular pH is important for membrane trafficking, but the determinants of pH homeostasis are poorly defined in plants. Electrogenic proton (H+) pumps depend on counter-ion fluxes to establish transmembrane pH gradients at the plasma membrane and endomembranes. Vacuolar-type H+-ATPase-mediated acidification of the trans-Golgi network (TGN) is crucial for secretion and membrane recycling. Pump and counter-ion fluxes are unlikely to fine-tune pH; rather, alkali cation/H+ antiporters, which can alter pH and/or cation homeostasis locally and transiently, are prime candidates. Plants have a large family of predicted cation/H+ exchangers (CHX) of obscure function, in addition to the well-studied K+(Na+)/H+ exchangers (NHX). Here, we review the regulation of cytosolic and vacuolar pH, highlighting the similarities and distinctions of NHX and CHX members. In planta, alkalinization of the TGN or vacuole by NHXs promotes membrane trafficking, endocytosis, cell expansion, and growth. CHXs localize to endomembranes and/or the plasma membrane, contribute to male fertility, pollen tube guidance, pollen wall construction, stomatal opening, and in soybean (Glycine max), tolerance to salt stress. Three-dimensional structural models and mutagenesis of Arabidopsis thaliana genes have allowed us to infer that AtCHX17 and AtNHX1 share a global architecture and a translocation core like bacterial Na+/H+ antiporters. Yet the presence of distinct residues suggests some CHXs differ from NHXs in pH sensing and electrogenicity. How H+ pumps, counter-ion fluxes, and cation/H+ antiporters are linked with signaling and membrane trafficking to remodel membranes and cell walls awaits further investigation.