Releasing the Cytokinin Brakes on Root Growth
Roots explore the soil for available water and nutrients, with deep roots providing water from the lower soil layers (Uga et al., 2015) and highly branched roots searching the soil for less-mobile nutrients such as phosphate, zinc, and manganese. Root architecture results from plant developmental programs, which are modified by environmental cues. The development of new lateral roots, which make up the majority of the root mass, depends on the plant hormone auxin. Auxin induces periclinal cell divisions that form lateral root founder cells (De Smet and Beeckman, 2011). Founder cells develop into lateral root primordia and lateral roots, a process modulated by other hormones, environmental signals, and developmental programs. Cytokinins create positional cues for new lateral roots, reducing lateral root formation and ensuring sufficient spacing between two neighboring lateral roots (Laplaze et al., 2007; Šimášková et al., 2015). However, the levels of root cytokinins under standard growth conditions are too high for optimal root growth, as increased root size might help the plant to absorb more water and improve nutrient acquisition.
The article by Ramireddy et al. (2018) published in this issue of Plant Physiology describes the effects of reducing cytokinin levels in roots on root growth. The authors expressed two Arabidopsis thaliana genes encoding the cytokinin-degrading enzymes cytokinin oxidase/dehydrogenase 1 and 2 (CKX1/2) in the barley (Hordeum vulgare) cv Golden Promise. Previous efforts to ectopically reduce cytokinin levels had detrimental effects on shoot growth (Pospíšilová et al., 2016), indicating the need for root-specific expression of these cytokinin catabolic enzymes. Using three different root-specific promotors from rice (Oryza sativa), Ramireddy et al. restricted the expression of CKX1/2 to the tissues involved in the formation of new lateral roots. The transgenic lines showed increased root surface area without any negative effects on shoot growth or yield of plants grown in well-watered conditions.
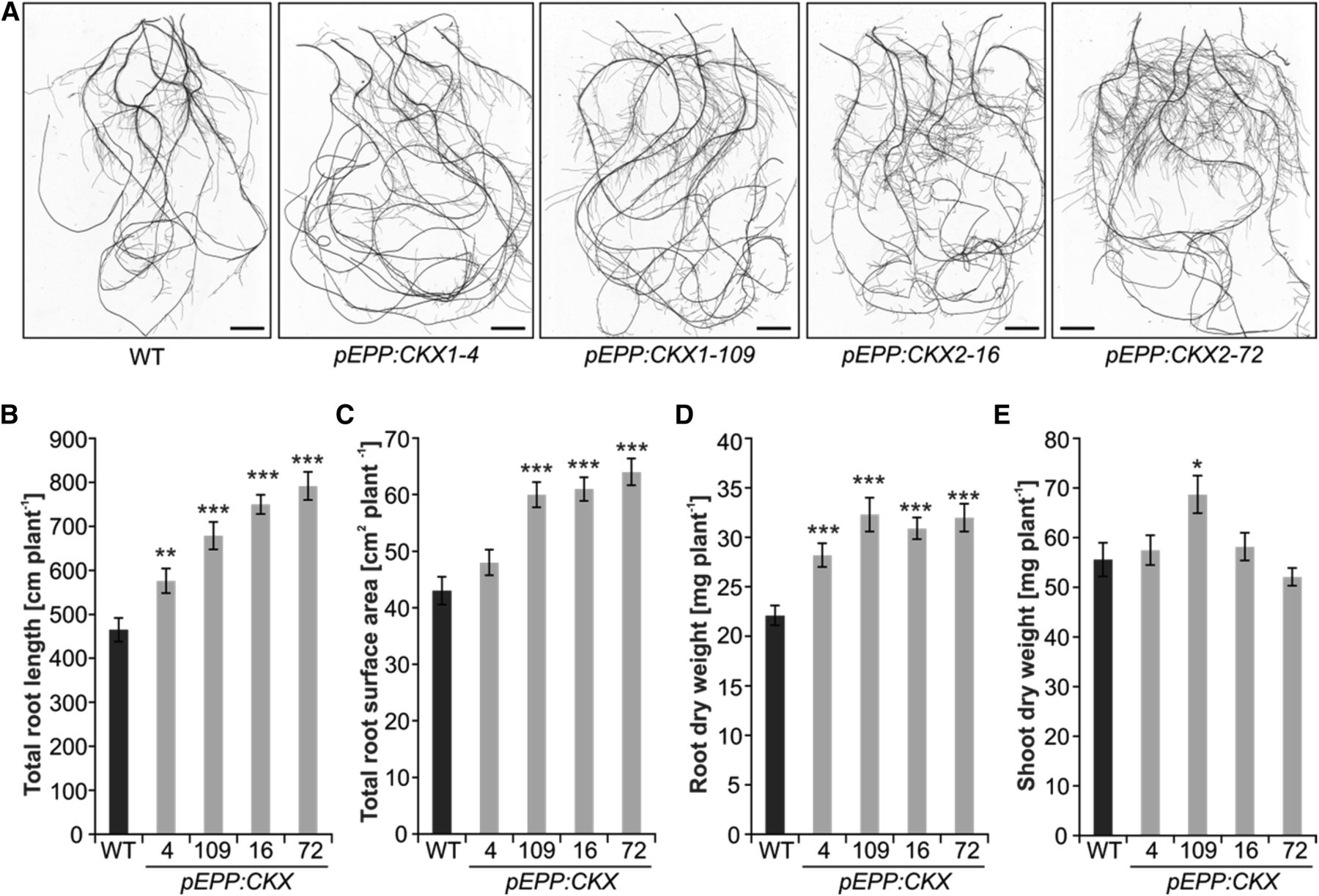
Transgenic plants with reduced cytokinin levels in the roots also showed increased drought tolerance, as the lines accumulated less abscisic acid (ABA) and Pro, had higher stomatal conductance, and showed improved carbon assimilation. In other studies, water deficit reduced production of shallow crown roots, and this reduction was more pronounced in wild, more drought-tolerant grass species (Sebastian et al., 2016). The reduced number of shallow crown roots likely redirected root growth into deeper soil layers containing more water. This direction of root growth toward water, also known as hydrotropism, was ascribed to ABA signaling (Dietrich et al., 2017). As cytokinin levels affect ABA responses (Tran et al., 2010), it will be important to establish whether the reduction of cytokinins in the roots affects the perception and translation of environmental signals crucial for effective soil exploration.
High root growth in low root cytokinin lines, as developed by Ramireddy et al., is beneficial under conditions where water is the only limiting factor. However, the root distribution in the soil rather than gross root mass might be more important in the field where phosphorous and nitrogen are scarce. Combining unleashed growth with root architecture and anatomy traits, such as cortical senescence (Schneider et al., 2017), will lower the carbon and nutrient costs of the root production, further improving the efficiency of soil exploration.
Large root systems allow the plant to explore more soil and to absorb more nutrients with low soil mobility (Lynch, 2018). High nutrient uptake ensures optimal plant growth and can add nutritional value when the minerals accumulate in the grains. The CKX lines developed by Ramireddy et al. accumulate more elements, such as sulfur and zinc, thereby increasing the nutritional value of the grain. The high accumulation of those nutrients might be additionally enhanced in the CKX lines, as cytokinin was observed to regulate the transporter genes for those ions (Brenner and Schmülling, 2012) and increase the number of passage cells in the Casparian strip (Andersen et al., 2018). Passage cells allow apoplastic transport of nutrients into the transpiration stream, but their increased number might also cause some leakage of the nutrients with high mobility, such as potassium. Mutants with reduced apoplastic barriers were previously observed to accumulate subsufficient potassium levels (Pfister et al., 2014), similar to the CKX lines developed by Ramireddy et al. The reduction of cytokinin in roots and increased soil foraging could be combined with other strategies, such as vacuolar ion storage (Connorton et al., 2017), for creating fortified crops.
The study by Ramireddy et al. is the first successful approach in monocot species to decrease cytokinin levels specifically in the roots, resulting in increased root growth and higher drought tolerance without yield penalty under standard growth conditions. Reducing the levels of cytokinins in the root additionally increases the nutritional value of barley by increasing zinc and sulfur accumulation. However, this high investment in roots might not benefit all crop systems, as high root mass in paddy-grown rice resulted in enhanced root transpiration, lower yield, and nutrient exudation, contributing to methanogenesis. In this context, lower root biomass, achieved by overexpression of the barley SUSIBA2 transcription factor, resulted in a shift of carbon flux, favoring the allocation of biomass to aboveground tissues, higher yield, and lower production of methane (Su et al., 2015). Before releasing the roots from the brakes on growth imposed by cytokinin, the agricultural system needs to be evaluated and the checkpoints imposed by cytokinins should be examined in the context of hydrotropism and effect on yield and nutrient use efficiency in heterogeneous field environments.
REFERENCES
Andersen TG, Naseer S, Ursache R, Wybouw B, Smet W, De Rybel B, Vermeer JEM, Geldner N (2018) Diffusible repression of cytokinin signalling produces endodermal symmetry and passage cells. Nature 555: 529–533
Brenner WG, Schmülling T (2012) Transcript profiling of cytokinin action in Arabidopsis roots and shoots discovers largely similar but also organ-specific responses. BMC Plant Biol 12: 112
Connorton JM, Jones ER, Rodríguez-Ramiro I, Fairweather-Tait S, Uauy C, Balk J (2017) Wheat vacuolar iron transporter TaVIT2 transports Fe and Mn and is effective for biofortification. Plant Physiol 174: 2434–2444
De Smet I, Beeckman T (2011) Asymmetric cell division in land plants and algae: the driving force for differentiation. Nat Rev Mol Cell Biol 12: 177–188
Dietrich D, Pang L, Kobayashi A, Fozard JA, Boudolf V, Bhosale R, Antoni R, Nguyen T, Hiratsuka S, Fujii N, et al. (2017) Root hydrotropism is controlled via a cortex-specific growth mechanism. Nat Plants3: 17057
Laplaze L, Benkova E, Casimiro I, Maes L, Vanneste S, Swarup R, Weijers D, Calvo V, Parizot B, Herrera-Rodriguez MB, et al. (2007) Cytokinins act directly on lateral root founder cells to inhibit root initiation. Plant Cell 19: 3889–3900
Lynch JP (2018) Rightsizing root phenotypes for drought resistance. J Exp Bot 69: 3279–3292
Pfister A, Barberon M, Alassimone J, Kalmbach L, Lee Y, Vermeer JEM, Yamazaki M, Li G, Maurel C, Takano J, et al. (2014) A receptor-like kinase mutant with absent endodermal diffusion barrier displays selective nutrient homeostasis defects. eLife 3: e03115
Pospíšilová H, Jiskrová E, Vojta P, Mrízová K, Kokáš F, Čudejková MM, Bergougnoux V, Plíhal O, Klimešová J, Novák O, et al. (2016) Transgenic barley overexpressing a cytokinin dehydrogenase gene shows greater tolerance to drought stress. N Biotechnol (5 Pt B) 33: 692–705
Schneider HM, Postma JA, Wojciechowski T, Kuppe C, Lynch JP (2017) Root cortical senescence improves growth under suboptimal availability of N, P, and K. Plant Physiol 174: 2333–2347
Sebastian J, Yee M-C, Goudinho Viana W, Rellán-Álvarez R, Feldman M, Priest HD, Trontin C, Lee T, Jiang H, Baxter I, et al. (2016) Grasses suppress shoot-borne roots to conserve water during drought. Proc Natl Acad Sci USA 113: 8861–8866
Šimášková M, O’Brien JA, Khan M, Van Noorden G, Ötvös K, Vieten A, De Clercq I, Van Haperen JMA, Cuesta C, Hoyerová K, et al. (2015) Cytokinin response factors regulate PIN-FORMED auxin transporters. Nat Commun 6: 8717
Su J, Hu C, Yan X, Jin Y, Chen Z, Guan Q, Wang Y, Zhong D, Jansson C, Wang F, et al. (2015) Expression of barley SUSIBA2 transcription factor yields high-starch low-methane rice. Nature 523: 602–606
Tran L-SP, Shinozaki K, Yamaguchi-Shinozaki K (2010) Role of cytokinin responsive two-component system in ABA and osmotic stress signalings. Plant Signal Behav 5: 148–150
Uga Y, Kitomi Y, Yamamoto E, Kanno N, Kawai S, Mizubayashi T, Fukuoka S (2015) A QTL for root growth angle on rice chromosome 7 is involved in the genetic pathway of DEEPER ROOTING 1. Rice (N Y) 8: 8







