Plant Physiology Focus Issue on Stomata published
The June 2017 issue of Plant Physiology is a Focus Issue on stomata; here is the editorial overview to this issue by Michael R. Blatt, Plant Physiology, Editor-in-Chief, and Tim J. Brodribb and Keiko U. Torii, Plant Physiology Editors.
Small Pores with a Big Impact
The guard cells surrounding stomatal pores of leaves are, at once, the best characterized and some of the most intriguing of cells in the plant kingdom. There are, of course, some very basic reasons for our interest in stomatal guard cells. The stomatal pore connects the inner air space of the leaf with the atmosphere, providing a route for gaseous exchange that bypasses the otherwise impermeable cuticle on the outer epidermal surface of land plants. Vascular plants regulate the pore aperture in response to an array of environmental and endogenous stimuli. Stomata thus play a critical role in enabling CO2 entry to the leaf for photosynthesis while protecting the hydrated environment needed by the photosynthetic tissues within. Stomata have a major influence on global water and carbon cycles, and they lie at the center of the crisis in water availability and crop production now expected to unfold over the next 20 to 30 years. Just how big an impact can such small pores have? As an example, we note that incorporating stomatal transpiration was a key factor behind much of the advance in atmospheric modeling of the 1990s, enabling weather prediction with the hour-by-hour precision that we recognize today.
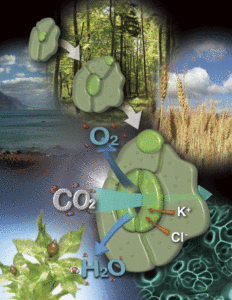 This Focus Issue on Stomata is the first dedicated to the evolution, development, and physiology of guard cells, their fundamental relevance to plants and to society. Several reviews and articles in this Focus Issue center around stomata as a target for genetic engineering to improve water use efficiency and their importance for crop survival. The Update reviews of Vialet-Chabrand et al. (2017b) and Matthews et al. (2017) address the potential for improved water use efficiencies by enhancing the kinetics of guard cell responses to environmental cues relevant for photosynthesis. Hughes et al. (2017) demonstrate the efficacy of manipulating stomatal densities to enhance water use efficiency and drought tolerance, while Hochberg et al. (2017) highlight the interplay between stomatal function, water delivery to the leaf, and leaf shedding in the field.
This Focus Issue on Stomata is the first dedicated to the evolution, development, and physiology of guard cells, their fundamental relevance to plants and to society. Several reviews and articles in this Focus Issue center around stomata as a target for genetic engineering to improve water use efficiency and their importance for crop survival. The Update reviews of Vialet-Chabrand et al. (2017b) and Matthews et al. (2017) address the potential for improved water use efficiencies by enhancing the kinetics of guard cell responses to environmental cues relevant for photosynthesis. Hughes et al. (2017) demonstrate the efficacy of manipulating stomatal densities to enhance water use efficiency and drought tolerance, while Hochberg et al. (2017) highlight the interplay between stomatal function, water delivery to the leaf, and leaf shedding in the field.
Guard cells attract much attention also as models for understanding evolution, development, and cellular physiology. At the heart of all biological research is a quest for understanding how life arises from what is more than the sum of its parts, in other words a search for what emerges from synergistic interactions between the respective components that underpin the biological process. The reviews and articles of this Focus Issue are testament to this, more fundamental interest in the guard cells, and they repeatedly highlight the emergent characteristics of life. The quest for these synergies—the nonlinearities within the biological mechanisms—is a theme that runs through research into stomatal guard cells at many levels.
Such nonlinearities are certainly evident in evolution. Stomata first appeared as simple pores on the surfaces of early land plants, evident in fossilized remains. These pores probably served to speed the desiccation of reproductive structures by contrast with the functions of stomata in vascular plants today. When did stomata evolve to moderate water loss? How did they acquire the molecular machinery that drives stomatal movements? And when was the complex network of signal cascades assembled to sensitize stomatal dynamics to environmental, hormonal, and pathogenic stimuli? The Update review by Franks et al. (2017) revisits these questions across temporal and spatial scales. Brodribb and McAdam (2017) highlight a selection of evolutionary boundaries and transitions in relation to signaling by the hormone abscisic acid. These boundaries remain open to debate, as noted by Hõrak et al. (2017) and by Cai et al. (2017). The questions surface again in the context of Crassulacean acid metabolism stomata in the Update review by Males and Griffiths (2017) and in the Correspondence from Bräutigam et al. (2017). McElwain and Steinthorsdottir (2017) as well as Renzaglia et al. (2017) approach these questions also from the perspective of fossil records. Renewed interest in the topic is certain to throw up new insights in the coming years.
Guard cells arise from the surrounding epidermal pavement cells as the leaf expands. Here, too, they present a model for cellular organization and differentiation that incorporates a wealth of nonlinearities, beginning with the asymmetry in cell division that gives rise to the guard mother cell. We dedicate this Focus Issue to the memory of Fred Sack (Bergmann et al., 2017), whose pioneering studies of mutants in stomatal development, as well as his infectious humor and generosity, formed the cornerstone of much research that has followed over the past two decades. The Update review by Chater et al. (2017) touches on some of the most recent developments on this topic. As an example of how this knowledge may be applied, again we point the reader to the research article by Hughes et al. (2017). de Marcos et al. (2017) uncover an unexpected interaction between brassinosteroid signaling and transcriptional control behind cell fates leading to stomatal development. Related articles that bridge the gap between the development of the stomatal cell wall and stomatal function (Huang et al., 2017; Hunt et al., 2017) will also be of interest in this context.
Of course, guard cells are widely recognized as the premier plant cell model for studies of membrane transport, signaling, and cellular homeostasis. Ion transport across the plasma membrane and tonoplast of the guard cell, and the water flux that follows, generates the turgor to drive guard cell volume and, hence, changes in stomatal aperture. In this Focus Issue, Jezek and Blatt (2017) review the most salient literature pertaining to transport. They stress the importance of quantitative physiological data and their application through dynamic modeling as tools to understand the synergies in transport and their emergent behaviors. Eisenach and De Angeli (2017) review in detail the substantial advances in knowledge of the vacuole and its role in guard cells. Inoue and Kinoshita (2017) discuss recent advances in dissecting the mechanisms behind light-mediated regulation of the H+-ATPases that energize the guard cell plasma membrane. The reviews by Males and Griffiths (2017) and Santelia and Lunn (2017) encapsulate the several exciting new developments that point to a complex coupling between carbohydrate metabolism and membrane transport in guard cells as well as its relationship to photosynthesis in the underlying mesophyll of leaves. Stomata are also points of access for pathogens to these delicate, nutrient-filled tissues. Melotto et al. (2017) review progress over the past decade in understanding how many pathogens commandeer guard cell signal cascades in order to enter the leaf tissues through the stomatal pore.
Accompanying research articles highlight several aspects of the signaling and regulatory networks affecting these processes. Malcheska et al. (2017) describe a potential role for sulfate, delivered via the xylem in poplar, as a signal that affects anion channel gating of the guard cells. Jalakas et al. (2017) revisit the role of the ERA1 farnesyl transferase to show that its actions extend well beyond abscisic acid signaling. Wang et al. (2017) address the interplay between small GTPase activities, the cytoskeleton, and phytochrome-mediated stomatal movements. Takahashi et al. (2017) examine hormonal signaling in the context of transcriptional controls that are important over longer timescales for the expression of selected ion channels in guard cells. Cerutti et al. (2017) describe the structure and physiology of hydathodes, another set of pores on the surface of the leaf with close access to the xylem, and the defense mechanisms they present to bacterial invasion.
Mathematical modeling has a long history in biology and is gaining pace as an approach to addressing the complexities of stomatal function. The Update reviews by Buckley (2017) and Franks et al. (2017) address this background, highlighting major gaps that remain to be bridged, notably between the microscopic and macroscopic scales of the guard cell and the plant canopy as well as between deterministic and global phenomenological approaches. Deans et al. (2017) refine a hydraulic model and propose a link to hormonal signaling in conifers. Finally, the research article of Vialet-Chabrand et al. (2017a) uses a well-established mathematical model of ion transport in stomatal guard cells (Chen et al., 2012; Hills et al., 2012), applying global parameter sensitivity analysis to identify potential targets for manipulation of stomatal kinetics.
In closing, we note that research over the past four decades has produced a vast wealth of information about guard cells, their development, membrane transport and its regulation, and the coordination of transport with metabolism. Indeed, our understanding of component processes of ion transport, especially, are sufficiently complete as to enable quantitative physiological models of stomatal guard cells with true predictive power, such as employed by Vialet-Chabrand et al. (2017a). Nonetheless, if we are to use our knowledge of stomata to improve crop resilience and agricultural capacity in marginal areas, then stomatal gas exchange from the leaf canopy must be linked to the mechanics of stomatal ion transport, metabolism, and their appearance during leaf expansion. Certainly, an understanding of the dynamics and coordination that lie behind stomatal formation and movements will be critical to a rational effort toward their engineering in the future.
REFERENCES
Bergmann D, Clare D, Samuels L, Kiss JZ (2017) A celebration of Fred David Sack. Plant Physiol 174: 470–472
Bräutigam A, Schlüter U, Eisenhut M, Gowik U (2017) On the evolutionary origin of CAM photosynthesis. Plant Physiol 174: 473–477
Brodribb TJ, McAdam SAM(2017) Evolution of the stomatal regulation of plant water content. Plant Physiol 174: 639–649
Buckley TN (2017) Modeling stomatal conductance. Plant Physiol 174: 572–582
Cai S, Chen G, Wang Y, Huang Y, Marchant DB, Wang Y, Yang Q, Dai F, Hills A, Franks PJ, et al. (2017) Evolutionary conservation of ABA signaling for stomatal closure. Plant Physiol 174: 732–747
Cerutti A, Jauneau A, Auriac M-C, Lauber E, Martinez Y, Chiarenza S, Leonhardt N, Berthomé R, Noël LD (2017) Immunity at cauliflower hydathodes controls systemic infection by Xanthomonas campestris pv campestris. Plant Physiol 174: 700–716
Chater CCC, Caine RS, Fleming AJ, Gray JE (2017) Origins and evolution of stomatal development. Plant Physiol 174: 624–638
Chen ZH, Hills A, Baetz U, Amtmann A, Lew VL, Blatt MR (2012) Systems dynamic modeling of the stomatal guard cell predicts emergent behaviors in transport, signaling, and volume control. Plant Physiol 159: 1235–1251
de Marcos A, Houbaert A, Triviño M, Delgado D, Martín-Trillo M, Russinova E, Fenoll C, Mena M (2017) A mutation in the bHLH domain of the SPCH transcription factor uncovers a BR-dependent mechanism for stomatal development. Plant Physiol 174: 823–842
Deans RM, Brodribb TJ, McAdam SAM (2017) An integrated hydraulic-hormonal model of conifer stomata predicts water stress dynamics. Plant Physiol 174: 478–486
Eisenach C, De Angeli A (2017) Ion transport at the vacuole during stomatal movements. Plant Physiol 174: 520–530
Franks PJ, Berry JA, Lombardozzi DL, Bonan GB (2017) Stomatal function across temporal and spatial scales: deep-time trends, land-atmosphere coupling, and global models. Plant Physiol 174: 583–602
Hills A, Chen ZH, Amtmann A, Blatt MR, Lew VL (2012) OnGuard, a computational platform for quantitative kinetic modeling of guard cell physiology. Plant Physiol 159: 1026–1042
Hochberg U, Windt CW, Ponomarenko A, Zhang Y-J, Gersony J, Rockwell FE, Holbrook NM (2017) Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems. Plant Physiol 174: 764–775
Hõrak H, Kollist H, Merilo E (2017) Fern stomatal responses to ABA and CO2 depend on species and growth conditions. Plant Physiol 174: 672–679
Huang Y-C, Wu H-C, Wang Y-D, Liu C-H, Lin C-C, Luo D-L, Jinn T-L (2017) PECTIN METHYLESTERASE34 contributes to heat tolerance through its role in promoting stomatal movement. Plant Physiol 174: 748–763
Hughes J, Hepworth C, Dutton C, Dunn JA, Hunt L, Stephens J, Waugh R, Cameron DD, Gray JE (2017) Reducing stomatal density in barley improves drought tolerance without impacting on yield. Plant Physiol 174: 776–787
Hunt L, Amsbury S, Baillie A, Movahedi M, Mitchell A, Afsharinafar M, Swarup K, Denyer T, Hobbs JK, Swarup R, et al. (2017) Formation of the stomatal outer cuticular ledge requires a guard cell wall proline-rich protein. Plant Physiol 174: 689–699
Inoue S-i, Kinoshita T (2017) Blue light regulation of stomatal opening and the plasma membrane H+-ATPase. Plant Physiol 174: 531–538
Jalakas P, Huang Y-C, Yeh Y-H, Zimmerli L, Merilo E, Kollist H, Brosche M (2017) The role of ENHANCED RESPONSES TO ABA1 (ERA1) in Arabidopsis stomatal responses is beyond ABA signaling. Plant Physiol 174: 665–671
Jezek M, Blatt MR (2017) The membrane transport system of the guard cell and its integration for stomatal dynamics. Plant Physiol 174: 487–519
Malcheska F, Ahmad A, Batool S, Müller HM, Ludwig-Müller J, Kreuzwieser J, Randewig D, Hänsch R, Mendel RR, Hell R, et al. (2017) Drought-enhanced xylem sap sulfate closes stomata by affecting ALMT12 and guard cell ABA synthesis. Plant Physiol 174: 798–814
Males J, Griffiths H (2017) Stomatal biology of CAM plants. Plant Physiol 174: 550–560
Matthews JSA, Vialet-Chabrand SRM, Lawson T (2017) Diurnal variation in gas exchange: the balance between carbon fixation and water loss. Plant Physiol 174: 614–623
McElwain JC, Steinthorsdottir M (2017) Paleoecology, ploidy, paleoatmospheric composition, and developmental biology: a review of the multiple uses of fossil stomata. Plant Physiol 174: 650–664
Melotto M, Zhang L, Oblessuc PR, He SY (2017) Stomatal defense a decade later. Plant Physiol 174: 561–571
Renzaglia KS, Villarreal JC, Piatkowski BT, Lucas JR, Merced A (2017) Hornwort stomata: architecture and fate shared with 400-million-year-old fossil plants without leaves. Plant Physiol 174: 788–797
Santelia D, Lunn JE (2017) Transitory starch metabolism in guard cells: unique features for a unique function. Plant Physiol 174: 539–549
Takahashi Y, Ebisu Y, Shimazaki K-I (2017) Reconstitution of abscisic acid signaling from the receptor to DNA via bHLH transcription factors. Plant Physiol 174: 815–822
Vialet-Chabrand S, Hills A, Wang Y, Griffiths H, Lew V, Lawson T, Blatt MR, Rogers S (2017a) Global sensitivity analysis of OnGuard models identifies key hubs for transport interaction in stomatal dynamics. Plant Physiol 174: 680–688
Vialet-Chabrand SRM, Matthews JSA, McAusland L, Blatt MR, Griffiths H, Lawson T (2017b) Temporal dynamics of stomatal behavior: modeling and implications for photosynthesis and water use. Plant Physiol 174: 603–613
Wang W, Liu Z, Bao L-J, Zhang S-S, Zhang C-G, Li X, Li H-X, Zhang X-L, Bones AM, Yang Z, Chen Y-L (2017) The RopGEF2-ROP7/ROP2 pathway activated by phyB suppresses red light-induced stomatal opening. Plant Physiol 174: 717–731


Leave a Reply
Want to join the discussion?Feel free to contribute!