New interacting partners of BLADE-ON-PETIOLE in regulation of plant development
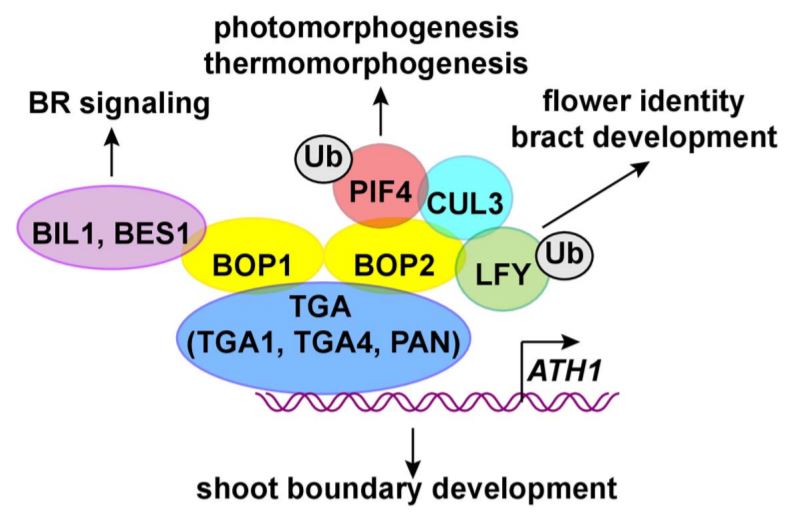
Organ boundaries are junctions that separate initiating lateral organs from the meristem or other plant parts. Cells at the boundary often exhibit slow growth rates and morphology distinct from that of the surrounding cells, and their development are controlled by complex gene networks. Lateral organ boundary marker genes BLADE-ON-PETIOLE1 (BOP1) and BOP2 are two closely related members in the BTB/POZ (Broad Complex, Tramtrack and Bric-a-brac/POX virus and Zinc finger) domain family, encoding plant-specific transcriptional co-activators. As indicated by the gene name, a dominant negative mutant of bop1 was first identified with ectopic leaf growth on petioles from an ethylmethane sulfonate mutant screen in Arabidopsis thaliana. Ectopic outgrowth also occurs in the stem and at the base of floral organs due to ectopic meristematic activities in these tissues (Ha et al., 2003). In addition, BOP1 and BOP2 function redundantly in various aspects of lateral organ and boundary development, including leaf, stem and flower patterning, and abscission zone development (Khan et al., 2014). BTB/POZ proteins lack a DNA binding domain and interact with TGACG-motif binding (TGA) class of basic leucine zipper (bZIP) transcription factors for DNA binding (Khan et al., 2014). Previous analysis showed that BOP1/2 physically interact with PERIANTHIA (Fig. 1), a class V TGA, to regulate flower development, particularly sepal numbers (Hepworth et al., 2005). In this issue of Plant Physiology, Wang et al. (2019) demonstrate that BOP1/2 and two clade I TGA proteins, TGA1 and TGA4, together regulate meristem maintenance and inflorescence architecture.
 Wang et al. (2019) use yeast two-hybrid and bimolecular fluorescence complementation assays to show that TGA1/4 and BOP1/2 physically interact. Promoter-GUS assays show that TGA1/4 and BOP1/2 are all referentially expressed at organ junctions, although TGA1/4 are also expressed in roots and vascular tissues of leaf and stem. tga1, tga4 or tga1 tga4 mutants do not show a bop1/2 phenotype; however, TGA1 and TGA4 are required for BOP2 promotion of stem elongation, as the short-stem phenotype in BOP2 overexpression lines is diminished in the tga1 tga2 mutant background. TGA1/4 and BOP1/2 also share the same upstream regulators of paralogous BELL homeodomain TFs, PENNYWISE (PNY) and POUND-FOOLISH (PNF), which regulate shoot apical meristem maintenance and inflorescence development (Wang et al., 2019). Moreover, chromatin immunoprecipitation assays revealed that both TGA1/4 and BOP1/2 associate with the same promoter region of ARABIDOPSIS THALIANA HOMEOBOX GENE1 (ATH1), which encodes another BELL homeodomain TF that regulates boundary development of shoot organs (Gómez-Mena and Sablowski, 2008). BOP1 promotion of ATH1 expression in an ATH1pro:GUS assay was abolished in the tga1 tga4 background, confirming the role of BOP1/2 and ATG1/4 as transcriptional co-activators of ATH1 expression (Fig. 1) (Wang et al., 2019).
Wang et al. (2019) use yeast two-hybrid and bimolecular fluorescence complementation assays to show that TGA1/4 and BOP1/2 physically interact. Promoter-GUS assays show that TGA1/4 and BOP1/2 are all referentially expressed at organ junctions, although TGA1/4 are also expressed in roots and vascular tissues of leaf and stem. tga1, tga4 or tga1 tga4 mutants do not show a bop1/2 phenotype; however, TGA1 and TGA4 are required for BOP2 promotion of stem elongation, as the short-stem phenotype in BOP2 overexpression lines is diminished in the tga1 tga2 mutant background. TGA1/4 and BOP1/2 also share the same upstream regulators of paralogous BELL homeodomain TFs, PENNYWISE (PNY) and POUND-FOOLISH (PNF), which regulate shoot apical meristem maintenance and inflorescence development (Wang et al., 2019). Moreover, chromatin immunoprecipitation assays revealed that both TGA1/4 and BOP1/2 associate with the same promoter region of ARABIDOPSIS THALIANA HOMEOBOX GENE1 (ATH1), which encodes another BELL homeodomain TF that regulates boundary development of shoot organs (Gómez-Mena and Sablowski, 2008). BOP1 promotion of ATH1 expression in an ATH1pro:GUS assay was abolished in the tga1 tga4 background, confirming the role of BOP1/2 and ATG1/4 as transcriptional co-activators of ATH1 expression (Fig. 1) (Wang et al., 2019).
The non-overlapping phenotypes of the bop1/2 and tga mutants indicate additional players in the BOP1/2 pathway. Indeed, BOP1 forms a complex with two master regulatory TFs in brassinosteroid (BR) signaling, BRZ-INSENSITIVE-LONG HYPOCOTYL 1/BRASSINAZOLE-RESISTANT 1 and BRI1-EMS-SUPPRESSOR 1, and inhibits their transport from cytosol to nucleus to suppress their activation of BR signaling (Shimada et al., 2015). BOP2 also interacts with the CULLIN3 ubiquitin ligase and recruits PHYTOCHROME INTERACTING FACTOR 4 (Zhang et al., 2017) or LEAFY TFs (Chahtane et al., 2018) as ubiquitination substrates for degradation, to modulate photomorphogenesis/thermomorphogenesis or flower identity, respectively (Fig. 1).
Taken together, these results suggest that BOP1/2 regulate lateral organ and boundary development by interacting with different partners to modulate gene transcription or mediate protein degradation. It would be interesting to study further how different interactors coordinate in BOP signaling.
Figure Legend
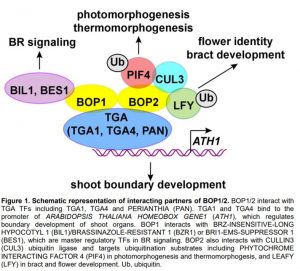
Figure 1. Schematic representation of interacting partners of BOP1/2. BOP1/2 interact with TGA TFs including TGA1, TGA4 and PERIANTHIA (PAN). TGA1 and TGA4 bind to the promoter of ARABIDOPSIS THALIANA HOMEOBOX GENE1 (ATH1), which regulates boundary development of shoot organs. BOP1 interacts with BRZ-INSENSITIVE-LONG HYPOCOTYL 1 (BIL1)/BRASSINAZOLE-RESISTANT 1 (BZR1) or BRI1-EMS-SUPPRESSOR 1 (BES1), which are master regulatory TFs in BR signaling. BOP2 also interacts with CULLIN3 (CUL3) ubiquitin ligase and targets ubiquitination substrates including PHYTOCHROME INTERACTING FACTOR 4 (PIF4) in photomorphogenesis and thermomorphogesis, and LEAFY (LFY) in bract and flower development. Ub, ubiquitin.
References
Chahtane H, Zhang B, Norberg M, LeMasson M, Thévenon E, Bakó L, Benlloch R, Holmlund M, Parcy F, Nilsson O (2018) LEAFY activity is post‐transcriptionally regulated by BLADE ON PETIOLE 2 and CULLIN 3 in Arabidopsis. New Phytologist 220: 579-592
Gómez-Mena C, Sablowski R (2008) ARABIDOPSIS THALIANA HOMEOBOX GENE1 establishes the basal boundaries of shoot organs and controls stem growth. The Plant Cell 20: 2059-2072
Ha CM, Kim G-T, Kim BC, Jun JH, Soh MS, Ueno Y, Machida Y, Tsukaya H, Nam HG (2003) The BLADE-ON-PETIOLE 1 gene controls leaf pattern formation through the modulation of meristematic activity in Arabidopsis. Development 130: 161-172
Hepworth SR, Zhang Y, McKim S, Li X, Haughn GW (2005) BLADE-ON-PETIOLE–dependent signaling controls leaf and floral patterning in Arabidopsis. The Plant Cell 17: 1434-1448
Khan M, Xu H, Hepworth SR (2014) BLADE-ON-PETIOLE genes: setting boundaries in development and defense. Plant Science 215: 157-171
Shimada S, Komatsu T, Yamagami A, Nakazawa M, Matsui M, Kawaide H, Natsume M, Osada H, Asami T, Nakano T (2015) Formation and dissociation of the BSS1 protein complex regulates plant development via brassinosteroid signaling. The Plant Cell 27: 375-390
Wang Y, SALASINI BC, Khan M, Devi B, Bush M, Subramaniam R, Hepworth SR (2019) Clade I TGA bZIP transcription factors mediate BLADE-ON-PETIOLE-dependent regulation of development. Plant Physiology: pp. 00805.02018
Zhang B, Holmlund M, Lorrain S, Norberg M, Bako L, Fankhauser C, Nilsson O (2017) BLADE-ON-PETIOLE proteins act in an E3 ubiquitin ligase complex to regulate PHYTOCHROME INTERACTING FACTOR 4 abundance. eLife 6: e26759