Evidence for Two Distinct Stages in Secondary Cell Wall Formation of Xylem
A hallmark of xylem development is the deposition of secondary cell wall material in specific patterns (reviewed in Patrick et al, 2007). These cell wall deposits structurally reinforce the xylem to withstand negative pressure during water transport and differ in different xylem cell types. While it is not entirely clear how the patterns of deposition are established, microtubule arrays assemble at the future sites of cell wall deposits, and cellulose synthase (CesA) complexes appear to move along the microtubule tracks (reviewed in Pesquet et al., 2011). CELLULOSE SYNTHASE INTERACTING PROTEIN 1 (CSI1), also known as POM-POM2 (POM2), interacts with CesAs and has been suggested to act as a scaffold mediating their movement along the microtubules (Gu and Somerville, 2010; Bringmann et al., 2012; Li et al., 2012). New work from Schneider et al. (2017), provides evidence that microtubules and CSI1/POM2 function early in secondary wall deposition, but are not required for later stages of secondary wall development.
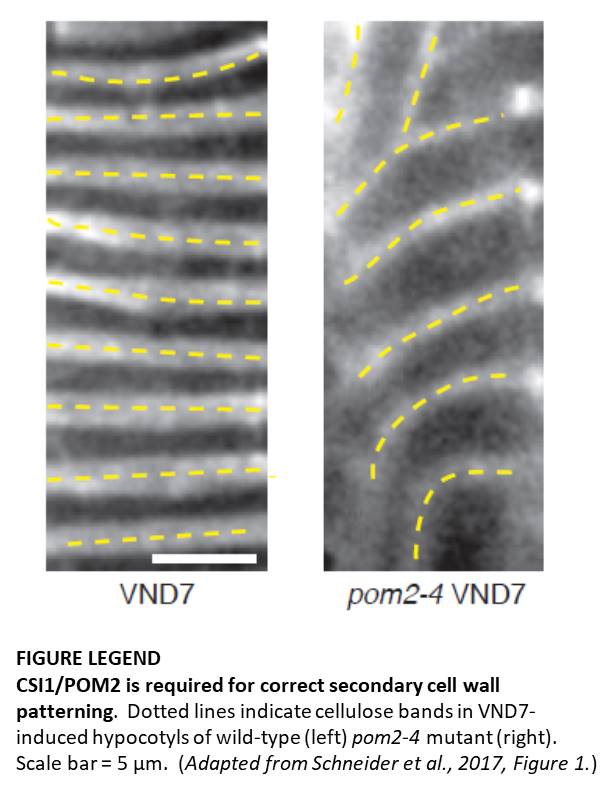 Schneider and coauthors explored the patterns of protoxylem vessel secondary walls in various model systems downregulated in CSI1/POM2. In contrast to the orderly array of cell wall bands in xylem vessels of wild-type Arabidopsis, pom2 and csi1 mutant plants had disordered secondary wall bands. Similarly, a VND7-inducible line (in which xylem differentiation can be induced in easily observable epidermis cells) in the wild-type background produced aligned bands of cell wall material. The VND7-inducible pom2 line still produced cell wall bands, but the spacing and alignment of the bands were less consistent (see Figure). These results indicate that CSI1/POM2 is not required for the formation of secondary wall deposits, but is involved in establishment of the pattern of their deposition.
Schneider and coauthors explored the patterns of protoxylem vessel secondary walls in various model systems downregulated in CSI1/POM2. In contrast to the orderly array of cell wall bands in xylem vessels of wild-type Arabidopsis, pom2 and csi1 mutant plants had disordered secondary wall bands. Similarly, a VND7-inducible line (in which xylem differentiation can be induced in easily observable epidermis cells) in the wild-type background produced aligned bands of cell wall material. The VND7-inducible pom2 line still produced cell wall bands, but the spacing and alignment of the bands were less consistent (see Figure). These results indicate that CSI1/POM2 is not required for the formation of secondary wall deposits, but is involved in establishment of the pattern of their deposition.
During primary wall synthesis, CSI1/POM2 moves with CesA complexes along microtubule tracks. Using a tagged version of CSI/POM2, Schneider et al. showed that CSI1/POM2 similarly moved along the bands during secondary wall synthesis, with movement patterns mirroring the rearrangement of the microtubule arrays. CSI1/POM2 moved at speeds similar to those of CesA complexes during secondary wall synthesis, supporting the relevance of the CSI/POM2 interaction with CesA complexes during this process.
In pom2 mutants, CesA movement and microtubules were misaligned during the early stages of secondary wall development. However, Schneider et al. provide evidence that the CesAs and microtubules become aligned later in development despite the absence of CSI1/POM2. In addition, treatment with oryzalin to disrupt microtubule arrays led to the absence of cell wall bands only when applied early during xylem vessel development: later treatment did not alter cell wall band deposition. Thus, it appears that microtubule arrays and CSI1/POM2 function in the establishment of the secondary wall deposition pattern, but that a different mechanism – which does not require CSI1/POM2 or microtubules – sustains later targeting of Golgi and secretory vesicles to the locations of deposition.
Schneider and coworkers have thus made nice use of a variety of analysis tools to provide experimental evidence for two complementary mechanisms at play in xylem wall development. The authors further showed that similar proteins could function in rice, supporting the idea that this is a general feature of secondary wall deposition. Many details of each phase of this process remain to be explored, including how the patterned microtubule arrays are established and how secretory compartments are targeted to the deposition sites during the second phase without requiring microtubules.
REFERENCES
Bringmann M., Li E., Sampathkumar A., Kocabek T., Hauser M.T., and Persson, S. (2012). POM-POM2/cellulose synthase interacting1 is essential for the functional association of cellulose synthase and microtubules in Arabidopsis. Plant Cell. 24: 163-177.
Gu, Y. and Somerville, C. (2010) Cellulose synthase interacting protein: a new factor in cellulose synthesis. Plant Signal Behav. 5: 1571-4.
Li, S., Lei, L., Somerville, C.R., and Gu, Y. (2012) Cellulose synthase interactive protein 1 (CSI1) links microtubules and cellulose synthase complexes. Proc Natl Acad Sci USA. 109: 185-90.
Pesquet, E., Korolev, A.V., Calder, G., and Lloyd, C.W. (2011) Mechanisms for shaping, orienting, positioning and patterning plant secondary cell walls. Plant Signal Behav. 6: 843-9.
Schneider, R., Tang, L., Lampugnani, E.R., Barkwill, S., Lathe, R., Zhang, Y. McFarlane, H.E., Pesquet, E., Niittyla, T., Mansfield, S.D., Zhou, Y., and Persson, S. (2017). Two Complementary Mechanisms Underpin Cell Wall Patterning during Xylem Vessel Development. Plant Cell. doi: 10.1105/tpc.17.00309.
Turner, S., Gallois, P., and Brown, D. (2007) Tracheary element differentiation. Annu Rev Plant Biol. 58: 407-33.