LAZY Genes Control Gravity Responses
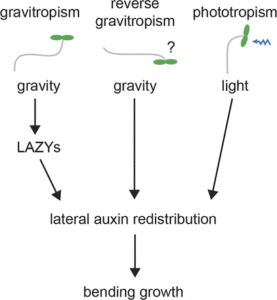 Plant architecture is shaped by innate developmental programs as well as by adaptive responses to environmental cues. For example, the primary growth axis of shoots and roots is vertical, with lateral branches adopting some characteristic angle with respect to the main axis. If the main axis is tipped from the vertical, gravitropism will bend the branch until it achieves its original orientation with respect to the gravity vector. Thus, a plant organ adopts a characteristic gravitational set point angle. Auxin gradients control the differential growth that sets the angle. Gravity-sensing leads to a relocalization of auxin transport proteins, which establishes an auxin gradient across the organ, followed by differential growth and bending. Investigations of the rice (Oryza sativa) lazy1 mutant, which displays a prostrate growth phenotype due to impaired gravitropism, led to the discovery of LAZY, a protein of unknown biochemical function that participates downstream of the gravity-sensing mechanism. Arabidopsis possesses six LAZY genes having spatially distinct expression patterns. By studying single and higher-order atlazy mutants, Yoshihara and Spalding (10.1104/pp.17.00942) have determined that AtLAZY1 is the principal determinant of inflorescence branch angle, whereas AtLAZY2 and AtLAZY4 determine lateral root branch angle. When the AtLAZY mechanism was disabled by multiple mutations, a separate mechanism for transducing the gravity vector into an auxin gradient was uncovered. This LAZY-independent mechanism created an oppositely oriented auxin gradient. Etiolated hypocotyls of the quadruple atlazy1,2,3,4 mutant were essentially agravitropic, but their phototropic response was robust. The authors propose that AtLAZY proteins control plant architecture by coupling gravity sensing to the formation of auxin gradients that override a LAZY-independent mechanism that creates an opposing gravity-induced auxin gradient.
Plant architecture is shaped by innate developmental programs as well as by adaptive responses to environmental cues. For example, the primary growth axis of shoots and roots is vertical, with lateral branches adopting some characteristic angle with respect to the main axis. If the main axis is tipped from the vertical, gravitropism will bend the branch until it achieves its original orientation with respect to the gravity vector. Thus, a plant organ adopts a characteristic gravitational set point angle. Auxin gradients control the differential growth that sets the angle. Gravity-sensing leads to a relocalization of auxin transport proteins, which establishes an auxin gradient across the organ, followed by differential growth and bending. Investigations of the rice (Oryza sativa) lazy1 mutant, which displays a prostrate growth phenotype due to impaired gravitropism, led to the discovery of LAZY, a protein of unknown biochemical function that participates downstream of the gravity-sensing mechanism. Arabidopsis possesses six LAZY genes having spatially distinct expression patterns. By studying single and higher-order atlazy mutants, Yoshihara and Spalding (10.1104/pp.17.00942) have determined that AtLAZY1 is the principal determinant of inflorescence branch angle, whereas AtLAZY2 and AtLAZY4 determine lateral root branch angle. When the AtLAZY mechanism was disabled by multiple mutations, a separate mechanism for transducing the gravity vector into an auxin gradient was uncovered. This LAZY-independent mechanism created an oppositely oriented auxin gradient. Etiolated hypocotyls of the quadruple atlazy1,2,3,4 mutant were essentially agravitropic, but their phototropic response was robust. The authors propose that AtLAZY proteins control plant architecture by coupling gravity sensing to the formation of auxin gradients that override a LAZY-independent mechanism that creates an opposing gravity-induced auxin gradient.







")