Trapped In the Rhizosheath: Root-Bacterial Interactions Modulate Ethylene Signaling
Author: Sjon Hartman 1,2
ORCID: 0000-0002-6709-6436
1 Plant Ecophysiology, Institute of Environmental Biology, Utrecht University, Padualaan 8, 3584 CH, Utrecht, The Netherlands
2 Corresponding author contact: [email protected]
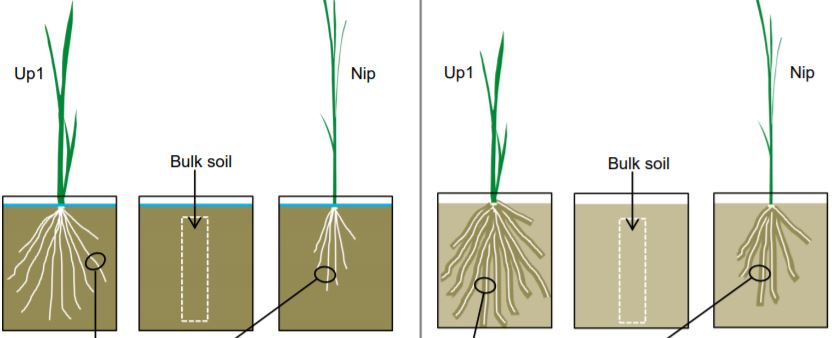
Even moderate droughts can greatly limit plant performance and reduce agricultural yield. Plants have evolved a wide range of traits to acclimate to drought (Yordanov et al., 2000), and enhancing these traits could improve drought tolerance in crops. One promising trait is the formation of the so-called ‘rhizosheath’ under drought. The rhizosheath is the soil that remains strongly attached to the root upon excavation. This protective layer enhances water and nutrient uptake by maintaining direct contact between the root and the soil (Brown et al., 2017). Rhizosheath formation occurs on the roots of many but not all angiosperms and is associated with several factors, including: root hairs, root and microbial mucilage, and the surrounding microorganisms (Brown et al., 2017). Rhizosheath production is enhanced by drought, but the mechanism by which this occurs remains elusive.
In this issue of Plant Physiology, Zhang et al. (2020) explore rhizosheath formation in rice (Oryza sativa L.). Using a transcriptomics approach, the authors found that moderate drought strongly enhanced ethylene signaling in the roots. Interestingly, ethylene is known to reduce primary root growth but promote root hair formation in many plant species (Dolan, 2001). Therefore, longer root hairs could contribute to rhizosheath formation under moderate drought stress by enhancing anchorage of the surrounding soil particles. The study by Zhang et al. (2020) opens up possibilities to unravel how this ethylene signal is generated and whether it is indeed required for rhizosheath establishment. Since ethylene is a gaseous phytohormone, ethylene signals are readily transmitted between cells, especially when the gas is trapped within plant tissues (Sasidharan et al., 2018). Due to restricted gas diffusion, this ethylene entrapment typically occurs in roots that experience flooding or high soil compaction (Hussain et al., 1999; Sasidharan et al., 2018). Interestingly, Zhang et al. (2020) show that ethylene signaling is increased in roots grown in drier soils compared to waterlogged roots. It is plausible that ethylene would also accumulate in rhizosheath-encased roots through reduced gas diffusion similarly to compacted soils (Hussain et al., 1999), but it would place ethylene signaling downstream of rhizosheath formation. Alternatively, ethylene signaling could be switched on in response to drought through crosstalk with other phytohormones or enhanced ethylene production (Sharp, 2002; Hopper et al., 2016). To follow up Zhang et al. (2020), future experiments could examine an ethylene insensitive rice mutant to address whether ethylene signaling during drought is indeed required for rhizosheath formation (Ma et al., 2013).
Zhang et al. (2020) also analyzed the bacterial composition of the rhizosheath and found that drought led to an enrichment of plant growth-promoting Enterobacteriaceae rhizobacteria. The authors isolated several bacterial strains and found that many Enterobacteriaceae bacteria showed high 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase activity. Since ACC is a precursor of ethylene, ACC deaminase-containing bacteria can reduce ethylene-mediated plant responses and promote growth during environmental stress (Ravanbakhsh et al., 2017). Interestingly, this high ACC deaminase activity of the rhizosheath Enterobacteriaceae bacteria appears to be at odds with the observed enhanced ethylene signaling in the roots. However, inoculation of dry soils with wild type Enterobacter aerogenes resulted in increased rhizosheath formation while inoculation with an ACC deaminase-deficient mutant strain did not (Zhang et al., 2020). The authors thereby confirmed that the ACC deaminase activity is indeed crucial for enhanced rhizosheath formation during drought.
How does bacterial-produced ACC deaminase contribute to enhanced rhizosheath formation and alleviate drought stress? One explanation could be that an initial ethylene signal upon drought is generated which increases root hair development and subsequent rhizosheath formation, but also temporarily limits primary root growth. Once this rhizosheath is established, the ACC deaminase-activity of the rhizosheath-colonized bacteria could lift the ethylene-impaired root growth and restore plant growth. Taken together, the results produced by Zhang et al. (2020) suggest that modulation of ethylene levels by root-colonizing bacteria contributes to enhanced rhizosheath formation during moderate drought stress. In the future it may be worthwhile to study the spatial and temporal dynamics of ACC and ethylene levels during rhizosheath formation, to uncover potential interaction effects. Revealing this interaction between a rhizosheath-forming plant species and its microbial partners could be pivotal in promoting rhizosheath formation in non-rhizosheath forming plants, with the aim to enhance drought tolerance.
LITERATURE CITED
Brown LK, George TS, Neugebauer K, White PJ (2017) The rhizosheath – a potential trait for future agricultural sustainability occurs in orders throughout the angiosperms. Plant Soil 418: 115–128
Dolan L (2001) The role of ethylene in root hair growth in Arabidopsis. J Plant Nutr Soil Sci 164: 141–145
Hopper DW, Ghan R, Schlauch KA, Cramer GR (2016) Transcriptomic network analyses of leaf dehydration responses identify highly connected ABA and ethylene signaling hubs in three grapevine species differing in drought tolerance. BMC Plant Biol 16: 1–20
Hussain A, Black CR, Taylor IB, Roberts JA (1999) Soil compaction. A role for ethylene in regulating leaf expansion and shoot growth in tomato? Plant Physiol 121: 1227–38
Ma B, He SJ, Duan KX, Yin CC, Chen H, Yang C, Xiong Q, Song QX, Lu X, Chen HW, Zhang WK, Lu TG, Chen SY, Zhang JS (2013) Identification of rice ethylene-response mutants and characterization of MHZ7/OsEIN2 in distinct ethylene response and yield trait regulation. Mol Plant 6: 1830–1848
Ravanbakhsh M, Sasidharan R, Voesenek LACJ, Kowalchuk GA, Jousset A (2017) ACC deaminase-producing rhizosphere bacteria modulate plant responses to flooding. J Ecol 105: 979–986
Sasidharan R, Hartman S, Liu Z, Martopawiro S, Sajeev N, van Veen H, Yeung E, Voesenek LACJ (2018) Signal Dynamics and Interactions during Flooding Stress. Plant Physiol 176: 1106–1117
Sharp RE (2002) Interaction with ethylene: Changing views on the role of abscisic acid in root and shoot growth responses to water stress. Plant, Cell Environ 25: 211–222
Yordanov I, Velikova V, Tsonev T (2000) Plant responses to drought, acclimation, and stress tolerance. Photosynthetica 38: 171–186