Tomatoes turn pale in the heat: high temperature reduces red and green pigmentation via phytochromes
The gardeners among us know that direct sunlight is a prerequisite to obtain full-sized, bright red tomato (Solanum lycopersicum) fruits. Indeed, light increases the accumulation of yellow and red carotenoid pigments, particularly lycopene, in ripe tomato fruits while it also boosts chlorophyll levels in the green parts of the plant. At the molecular level, accumulation of these pigments is regulated by multiple light signalling components, in particular the red/far-red light sensing phytochrome (phy) photoreceptors and their immediate downstream targets, the PHYTOCHROME INTERACTING FACTORs (PIFs) (Bianchetti et al., 2018; Gramegna et al., 2019; Rosado et al., 2019). Phytochromes act as a red/far-red light switch: upon red light perception, the inactive, red-light absorbing Pr form is converted into the biologically active Pfr form, which can in turn be inactivated by the absorption of far-red light. Active phytochromes elicit downstream responses in part by inhibiting and promoting the degradation of PIF transcription factors, thereby controlling gene expression in a light-dependent manner (Legris et al., 2017).
In Arabidopsis thaliana, phyB has been found to act not only as a photoreceptor, but also as a temperature sensor. In addition to far-red light-induced photoconversion, Pfr can be inactivated by a relaxation process termed thermal reversion as it is directly dependent on temperature; high ambient temperature speeds up this process and reduces phytochrome activity (Jung et al., 2016; Legris et al., 2016). Thermal reversion – or dark reversion, as it was originally called – has been observed for phytochromes of multiple species (Klose et al., 2020), but their control of developmental and physiological processes in response to ambient temperature has almost exclusively been investigated in Arabidopsis. In this issue of Plant Physiology, Bianchetti et al. (2020) show that phytochromes exert temperature-related functions in tomato, where they mediate the reduction in pigment accumulation at elevated temperatures
Given the function of phyB as a thermosensor in Arabidopsis, the authors decided to investigate a putative role for tomato phytochromes phyB1 and phyB2 in temperature-regulated processes. While elevated temperature reduced stem diameter, leaf area and branching, these responses where still present in a phyB1B2 double mutant, suggesting that the respective developmental processes are not controlled by phyB-mediated temperature sensing. Since phytochromes are known to affect pigment accumulation in tomato, the authors then decided to focus their investigation on metabolic traits. Elevated temperature indeed reduced leaf chlorophyll and carotenoid content, which is accompanied by impaired chloroplast biogenesis and differentiation as well as by alteration in the chloroplast’s ultrastructure. These changes are at least partially mediated by a reduction of phytochrome activity since a phyB1B2 double mutant showed constitutive reduction in pigment accumulation and chloroplast formation even when grown at low temperature. This observation is further corroborated by transcriptomic analyses, which revealed that heat reduces expression of carotenoid biosynthesis genes while it promotes expression of chlorophyll catabolic genes, both in a phyB1/B2-dependent manner (Figure 1).
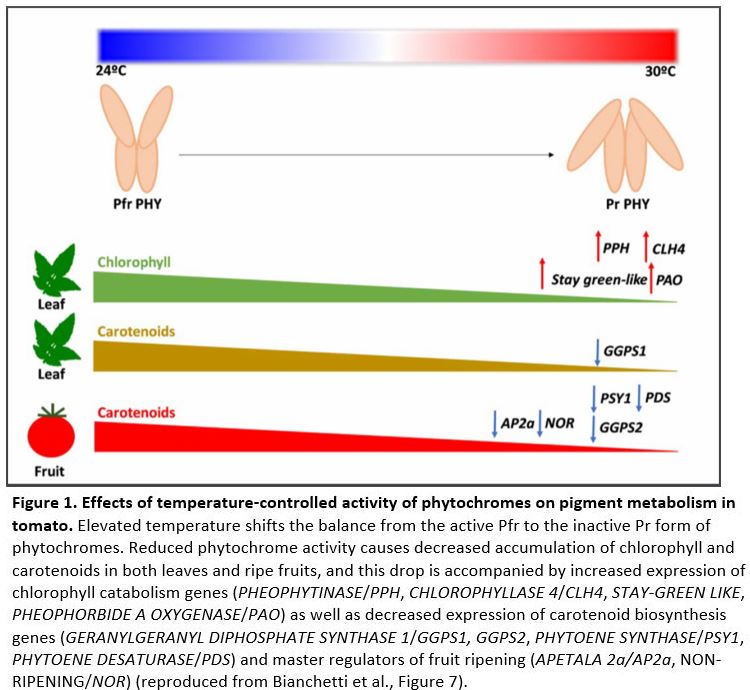 When Bianchetti et al. investigated ripe tomato fruits, they found that levels of the carotenoid lycopene and its precursors specifically decreased in response to elevated temperature. Previous results showed that phyA and phyB2 are core regulators of lycopene biosynthesis in tomato fruits (Bianchetti et al., 2018), and indeed, lycopene accumulation was constitutively reduced in a phyAB1B2 triple mutant as well as in PHYA and PHYB2 RNA interference (RNAi) lines that knock down the respective PHY genes selectively in fruits. Lycopene levels also correlated with the expression of lycopene biosynthesis genes in these genotypes, as well as with the expression of several master transcription factors known to control fruit ripening (Figure 1).
When Bianchetti et al. investigated ripe tomato fruits, they found that levels of the carotenoid lycopene and its precursors specifically decreased in response to elevated temperature. Previous results showed that phyA and phyB2 are core regulators of lycopene biosynthesis in tomato fruits (Bianchetti et al., 2018), and indeed, lycopene accumulation was constitutively reduced in a phyAB1B2 triple mutant as well as in PHYA and PHYB2 RNA interference (RNAi) lines that knock down the respective PHY genes selectively in fruits. Lycopene levels also correlated with the expression of lycopene biosynthesis genes in these genotypes, as well as with the expression of several master transcription factors known to control fruit ripening (Figure 1).
Taken together, the results obtained by Bianchetti et al. show that in tomato, phytochromes are key factors in metabolic responses to elevated temperature, suggesting that the phytochromes’ temperature-sensing capacity is of physiological relevance in species other than Arabidopsis. A recent study showed that tomato PIF4 also controls temperature responses, similarly to its Arabidopsis counterpart (Rosado et al., 2019). It thus appears that the light- and temperature-sensing phytochrome-PIF module is conserved in tomato, highlighting the crucial role these proteins play in sensing and reacting to the plant’s environment. Nevertheless, further investigation will be required to decipher the precise mechanisms of this regulatory module in tomato and other agriculturally-relevant species and to reveal commonalities and differences when compared to the canonical Arabidopsis phytochrome signalling pathway.
Martin Balcerowicz
Assistant Features Editor
Sainsbury Laboratory, University of Cambridge, Cambridge CB2 1LR, United Kingdom
Literature Cited
Bianchetti RE, Silvestre Lira B, Santos Monteiro S, Demarco D, Purgatto E, Rothan C, Rossi M, Freschi L (2018) Fruit-localized phytochromes regulate plastid biogenesis, starch synthesis, and carotenoid metabolism in tomato. J Exp Bot 69: 3573–3586
Gramegna G, Rosado D, Carranza APS, Cruz AB, Simon‐Moya M, Llorente B, Rodríguez‐Concepcíon M, Freschi L, Rossi M (2019) PHYTOCHROME-INTERACTING FACTOR 3 mediates light-dependent induction of tocopherol biosynthesis during tomato fruit ripening. Plant, Cell & Environment 42: 1328–1339
Jung J-H, Domijan M, Klose C, Biswas S, Ezer D, Gao M, Khattak AK, Box MS, Charoensawan V, Cortijo S, et al (2016) Phytochromes function as thermosensors in Arabidopsis. Science 354: 886–889
Klose C, Nagy F, Schäfer E (2020) Thermal Reversion of Plant Phytochromes. Molecular Plant 13: 386–397
Legris M, Klose C, Burgie ES, Rojas CCR, Neme M, Hiltbrunner A, Wigge PA, Schäfer E, Vierstra RD, Casal JJ (2016) Phytochrome B integrates light and temperature signals in Arabidopsis. Science 354: 897–900
Legris M, Nieto C, Sellaro R, Prat S, Casal JJ (2017) Perception and signalling of light and temperature cues in plants. The Plant Journal 90: 683–697
Rosado D, Trench B, Bianchetti R, Zuccarelli R, Alves FRR, Purgatto E, Floh EIS, Nogueira FTS, Freschi L, Rossi M (2019) Downregulation of PHYTOCHROME-INTERACTING FACTOR 4 Influences Plant Development and Fruit Production. Plant Physiology 181: 1360–1370






