The Hidden World of Mycorrhizae and Their Impact on Plant Health and Ecosystems
The trendy invisible world beneath our feet
Although the fungal world popped up in recent years thanks to several factors, including the Netflix documentary “Fantastic Fungi”, they have been on Earth for hundreds of millions of years, playing essential roles in ecosystems long before humans even existed. When we think about Robin Hood hiding in Sherwood Forest, we usually imagine trees, bushes, and dense vegetation. But forests are much more complex than what we see on the surface: beneath the soil, an enormous underground network connects roots, fungi, microbes, water, and nutrients in ways we are only beginning to understand. While we can see leaves, flowers, and tree canopies above ground, much of the activity that sustains plant life happens hidden below our feet.
At the center of this complex network are mycorrhizae, which, basically, are intimate symbiotic associations between plant roots and fungi that have shaped for more than 400 million years. The fungal networks extend far beyond individual roots, exploring the soil with microscopic filaments that transport nutrients and water across huge distances. For a very long time, plants have been seen mainly as independent organisms competing for limited resources. However, research increasingly reveals plants as part of highly interconnected underground systems in which fungi play central roles (Oyarte et al., 2025).
But what are mycorrhizae exactly, and why are they so important?
What are mycorrhizae?
The term mycorrhiza comes from the Greek words mykes, meaning fungus, and rhiza, meaning root. The combination of the two literally means ‘fungal root’. These symbiotic associations occur when fungi colonize plant roots and establish a interaction with their host (Janerertte, 1991). This interaction is maintained by two factors. On the one hand, plants supply the fungi with carbon compounds produced through photosynthesis, and on the other hand, the fungi help the plants to obtain nutrients and water from the soil. Mycorrhizal hyphae are thread-like structures that form extensive underground networks, acting as natural extensions of the root system and reaching regions of the soil that are inaccessible to the roots on their own. This association is very widespread. Between 80% and 90% of terrestrial plant species form mycorrhizal associations, highlighting their enormous ecological and evolutionary importance. In many ecosystems, plants rely heavily on these fungal partners for their survival and growth (Smith & Read, 2008).
The major types of mycorrhizas
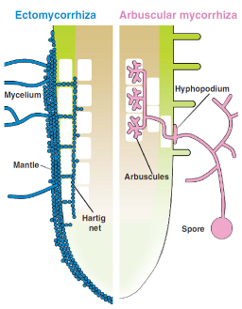
Illustration of root colonization structures in ectomycorrhizal (blue) and arbuscular mycorrhizal (pink) interactions. From Bonfante & Genre, 2010.
Not all mycorrhizae are the same. Although there are various forms of mycorrhizal associations, two main types predominate in most terrestrial ecosystems: arbuscular mycorrhizae and ectomycorrhizae. Arbuscular mycorrhizal fungi are by far the most widespread. They penetrate the cortical cells of the roots and form highly branched structures called arbuscules, which function as nutrient exchange points between the fungus and the plant. These associations are especially common in grasses, crops, and herbaceous plants. In contrast, ectomycorrhizae follow a different strategy. They surround the roots with a dense fungal sheath and grow between the root cells rather than penetrating them directly. These symbioses are characteristic of many forest trees, such as pines, oaks, birches, and beeches. Beyond these two groups, other mycorrhizal associations exist, such as orchid mycorrhizae and ericoid mycorrhizae, revealing the extraordinary diversity of interactions between plants and fungi that have evolved in different environments (Smith & Read, 2008).
The mechanics of the interaction
All successful interactions need good communication between both parties. In the case of mycorrhiza and plant roots, they start a complex molecular conversation in the soil, even before establishing any physical contact. Plant roots release compounds that stimulate fungal growth and, in turn, fungi generate signaling molecules that prepare the plant for colonization. Once they establish the symbiotic relationship, an exchange of resources takes place between the plant and the fungus. Fungi deliver nutrients such as phosphorus and nitrogen, as well as water and micronutrients, whereas plants allocate a substantial portion of the carbon fixed through photosynthesis to the fungi. However, this relationship is not as simple as an exchange of resources. The advantages and disadvantages of the interaction for each party might change depending on the environment, since some factors such as soil conditions, abiotic stresses, and the identity of both species can influence the beneficial interaction (Huey et al., 2020, Ho-Plágaro & García-Garrido, 2022).
Beyond nutrition
For many years, mycorrhizae have been studied for their role in helping plants to obtain nutrients, such as phosphate, but, although this remains one of their most important functions, research in the last few years has revealed that they can influence many other aspects of plant health. One of the best-known examples is their role in helping plants cope with biotic and abiotic stresses. Mycorrhizae can improve drought stress tolerance by enhancing water uptake and modulating the plant’s physiological responses, which helps them maintain growth under drought conditions. (Cosme, 2023). Fungi can also reduce the impact of soil pathogens by priming plant defenses and shaping the root-associated microbial communities (Umer et al., 2025). The effects of mycorrhizae are not limited only to individual plants. Their networks of hyphae contribute to soil aggregation and help improve soil structure. By binding soil particles together, fungal hyphae can increase soil stability and water retention, creating conditions that are beneficial for entire plant communities (Rillig & Mummey, 2006). These findings have changed the way we study mycorrhizae. Instead of acting simply as nutrient suppliers, mycorrhizae are increasingly recognized as key elements for plant health and the functioning of ecosystems. As climate change and soil degradation continue to threaten natural and agricultural ecosystems, understanding these relationships among fungi and plants may become more important than ever.
Ecosystem impacts and the “Mother Tree Hypothesis”
 While the partnership between a single plant and its fungus is intimate, the consequences play out at a far larger scale. Because a single fungal genotype can colonize the roots of many neighbouring plants, mycorrhizal hyphae can physically link individuals into what are called common mycorrhizal networks (CMNs). The existence of these networks is not in doubt; what they actually do for the plants they connect has become one of the more instructive controversies in plant science.
While the partnership between a single plant and its fungus is intimate, the consequences play out at a far larger scale. Because a single fungal genotype can colonize the roots of many neighbouring plants, mycorrhizal hyphae can physically link individuals into what are called common mycorrhizal networks (CMNs). The existence of these networks is not in doubt; what they actually do for the plants they connect has become one of the more instructive controversies in plant science.
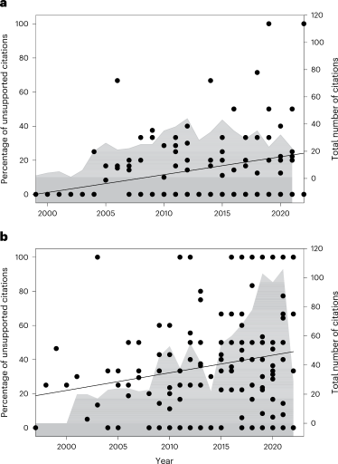
The popular version of the story is seductive. In it, “mother trees” recognize their own seedlings and route carbon, nitrogen, and even chemical warnings to them through a fungal “wood-wide web.” It is a tidy narrative, and it has been repeated everywhere from documentaries to bestselling books. But a careful reading of the primary literature tells a more sober story. In an influential 2023 perspective, Karst and colleagues evaluated three of the most widely repeated claims and found them to be running well ahead of the evidence (Karst et al., 2023). They concluded that the claims that CMNs are widespread in forests and that they transfer resources in ways that measurably improve seedling performance are insufficiently supported, because field results vary too widely and often admit simpler explanations. The most cinematic claim of all, that mature trees preferentially direct resources and defence signals to their own offspring through these networks, had, at the time of their review, no peer-reviewed published evidence to support it (Karst et al., 2023). The authors also documented a citation bias in the field: positive results were cited far more often than the more equivocal findings that accompanied them, which is how an idea can come to feel established long before the data justify it.
None of this means the underground is inert. It means the honest position is a humbler one: hyphae do move carbon and nutrients through soil, the direction and ecological significance of those movements are genuinely difficult to demonstrate, and a hypha acting as a passive conduit for diffusing molecules is not the same thing as a tree deliberately “talking” to its kin. 
Where the ecosystem-scale effects of mycorrhizas rest on firmer ground is in the soil itself. Fungal hyphae enmesh and bind soil particles, promoting the formation of stable aggregates that improve soil structure, porosity, and water retention (Rillig & Mummey, 2006). This physical engineering benefits whole plant communities rather than single hosts, and it feeds into one of the most striking recent findings about these fungi: their role in the global carbon cycle. In a 2023 meta-analysis of nearly 200 datasets, Hawkins and colleagues estimated that terrestrial plants allocate on the order of 13 gigatonnes of CO₂-equivalent of photosynthetically fixed carbon to mycorrhizal fungi each year (Hawkins et al., 2023). That figure is genuinely large, and it underscores how central these fungi are as an entry point for carbon into soil. It deserves one important caveat, however, one that the authors themselves emphasize: this is a measure of carbon allocated to the fungi, not carbon permanently locked away. How long carbon resides belowground and how much is respired, decomposed, stabilized on mineral surfaces, or returned to plants remains an open question (Hawkins et al., 2023). The soil is a major carbon pool, but treating an annual flux as if it were durable storage is precisely the kind of overstatement the field is learning to avoid.
Threats and disruptions
These hidden partnerships are not indestructible, and the most pervasive threats to them are human. Agriculture is the clearest case. The practices that define intensive farming, such as frequent tillage, heavy mineral fertilization, and low crop diversity, are also the practices most corrosive to mycorrhizal communities (Bowles et al., 2017). The mechanisms are intuitive. Ploughing physically shreds the mycelium outside of roots, severing the foraging networks the fungi depend on and that take time to rebuild. Abundant synthetic phosphorus and nitrogen remove much of the plant’s incentive to trade carbon for fungal nutrient delivery, weakening the symbiosis at its economic root (Johnson et al., 2008). And monocultures present the fungal community with a narrow, uniform set of hosts, which over time can select for a handful of stress-tolerant generalists at the expense of overall diversity (Guzman et al., 2021).
The magnitude of these effects can be considerable. A metaanalysis of tillage and cover cropping studies spanning five continents found that reducing soil disturbance and maintaining living roots through winter cover crops each increased mycorrhizal colonization of cash crops by roughly 30% (Bowles et al., 2017), a reminder that the losses are real but, encouragingly, often reversible. Land conversion can be far more severe: in tropical mountain ecosystems, the conversion of uncultivated land to agriculture has been associated with reductions in arbuscular mycorrhizal fungal richness of around 80% on average, with uncultivated soils harbouring roughly threefold more unique fungal species than their cultivated counterparts (Stewart et al., 2026). Beyond agriculture, the broader drivers of global change (nitrogen deposition, soil degradation, and shifting climate) apply additional, compounding pressure on communities that, being invisible and slow to recover, rarely figure into conservation planning at all.
Applications in agriculture and future research directions
The flip side of this vulnerability is opportunity. If conventional intensification degrades mycorrhizal function, then managing it offers a route toward more sustainable agriculture. The same evidence that documents the damage also points to the remedies: reduced or no-till systems, cover cropping, diversified rotations, and more judicious fertilizer use all tend to preserve or restore fungal colonization and diversity (Guzman et al., 2021, Bowles et al., 2017). The appeal is straightforward, as a healthy mycorrhizal community can improve a crop’s acquisition of phosphorus and water, buffer it against drought, and contribute to defence against soil-borne pathogens, reducing reliance on the very inputs whose overuse undermines the symbiosis in the first place.
Direct inoculation of crops with selected fungal strains is the more interventionist version of this idea, and it is already a commercial reality. Here, though, the science isn’t necessarily clean. The benefits of mycorrhizal symbiosis are strongly context dependent, varying with soil chemistry, climate, and the particular pairing of plant and fungal partners, so an inoculant that performs well in one field may do little in another. Introduced strains must also establish themselves against an existing native community. The most valuable research directions therefore, lie in understanding which fungal partners deliver which benefits under which conditions, how introduced and resident fungi interact, and how field management can reliably steer outcomes. Closing those gaps is what will determine whether mycorrhizas become a dependable tool of agronomy rather than an inconsistent one.
Conclusion
For most of the history of plant biology, the plant ended at the root tip. The study of mycorrhizas has redrawn that boundary, revealing that the great majority of land plants live in intimate partnership with fungi whose influence radiates outward from the individual root to the entire ecosystem. At the smallest scale, these fungi feed their hosts, ferrying phosphorus, nitrogen, and water that roots alone could never reach, and they harden those hosts against drought and disease. At a larger scale, their hyphae knit the soil itself together, building the stable structure on which whole plant communities depend. And at the largest scale of all, they stand at one of the main gateways through which atmospheric carbon enters the ground, handling a flux of photosynthetic products measured in gigatonnes each year. A single symbiosis, scaled across most of the world’s plants, becomes a force shaping global biogeochemistry.
That reach is also what makes their decline consequential. When tillage, heavy fertilization, and monoculture erode mycorrhizal communities, the losses are not confined to a single field; they ripple through soil stability, crop resilience, and the carbon cycle alike. The encouraging counterpart is that the same leverage works in reverse: by managing land in ways that sustain these fungi, dividends result across all those scales at once. Recognizing the full extent of what they do, and protecting their capacity to keep doing it, is among the more powerful and least visible opportunities we have for keeping ecosystems and the agriculture we draw from them healthy.
References:
Bowles, T. M., Jackson, L. E., Loeher, M., & Cavagnaro, T. R. (2017). ‘Ecological intensification and arbuscular mycorrhizas: a meta-analysis of tillage and cover crop effects,’ Journal of Applied Ecology, 54(6), 1785–1793. Available at: https://doi.org/10.1111/1365-2664.12815
Cosme, M. (2023). ‘Mycorrhizas drive the evolution of plant adaptation to drought,’ Communications Biology, 6(1), 346. Available at: https://doi.org/10.1038/s42003-023-04722-4
Guzman, A., Montes, M., Hutchins, L., DeLaCerda, G., Yang, P., Kakouridis, A., Dahlquist-Willard, R. M., Firestone, M. K., Bowles, T., & Kremen, C. (2021). ‘Crop diversity enriches arbuscular mycorrhizal fungal communities in an intensive agricultural landscape,’ New Phytologist, 231(1), 447–459. Available at: https://doi.org/10.1111/nph.17306
Hawkins, H.-J., Cargill, R. I. M., Van Nuland, M. E., Hagen, S. C., Field, K. J., Sheldrake, M., Soudzilovskaia, N. A., & Kiers, E. T. (2023). ‘Mycorrhizal mycelium as a global carbon pool,’ Current Biology, 33(11), R560–R573. Available at: https://doi.org/10.1016/j.cub.2023.02.027
Ho-Plágaro, T., & García-Garrido, J. M. (2022). ‘Molecular regulation of arbuscular mycorrhizal symbiosis,’ International Journal of Molecular Sciences, 23(11), 5960. Available at: https://doi.org/10.3390/ijms23115960
Huey, C. J., Gopinath, S. C. B., Uda, M. N. A., Zulhaimi, H. I., Jaafar, M. N., Kasim, F. H., Yaakub, A. R. W. (2020). ‘Mycorrhiza: a natural resource assists plant growth under varied soil conditions,’ 3 Biotech, 10(5), 204. Available at: https://doi.org/10.1007/s13205-020-02188-3
Janerette, C. A. (1991). ‘An introduction to mycorrhizae,’ The American Biology Teacher, 13-19. Available at: https://doi.org/10.2307/4449205
Johnson, N. C., Graham, J-H., Smith, F. A. ‘Functioning of mycorrhizal associations along the mutualism–parasitism continuum,’ New Phytologist, 135(4). Available at https://doi.org/10.1046/j.1469-8137.1997.00729.x
Karst, J., Jones, M. D., & Hoeksema, J. D. (2023). ‘Positive citation bias and overinterpreted results lead to misinformation on common mycorrhizal networks in forests,’ Nature Ecology & Evolution, 7(4), 501–511. Available at: 10.1038/s41559-023-01986-1
Oyarte Galvez, L., Bisot, C., Bourrianne, P., Cargill, R., Klein, M., van Son, M., van Krugten, J., Caldas, V., Clerc, T., Lin, K. K., Kahane, F., van Staalduine, S., Stewart, J. D., Terry, V., Turcu, B., van Otterdijk, S., Babu, A., Kamp, M., Seynen, M., Steenbeek, B., Zomerdijk, J., Tutucci, E., Sheldrake, M., Godin, C., Kokkoris, V., Stone, H. A., Kiers, E. T., Shimizu, T. S. (2025). ‘A travelling-wave strategy for plant–fungal trade,’ Nature, 639(8053), 172-180. Available at: https://doi.org/10.1038/s41586-025-08614-x
Rillig, M. C., & Mummey, D. L. (2006). ‘Mycorrhizas and soil structure,’ New Phytologist, 171(1), 41–53. Available at: https://doi.org/10.1111/j.1469-8137.2006.01750.x
Smith, S. E., Read, D. (2008). ‘Mycorrhizal symbiosis,’ Academic Press. Available at: https://doi.org/10.1016/B978-0-12-370526-6.X5001-6
Stewart, J. D., Ramirez, D. X., Leon-Reyes, A., Barriga, N., Llerena, S., Manley, B. F., Carpintero-Salvador, N., Ruiz-Uriguen, M., Raaijmakers, J. M., Kiers, E. T., Weedon, J. T. (2026). ‘Land use change reshapes climate-driven diversity patterns of tropical arbuscular mycorrhizal fungi,’ Molecular Ecology. Available at: https://doi.org/10.1111/mec.70253
Umer, M., Anwar, N., Mubeen, M., Li, Y., Ali, A., Alshaharni, M. O., & Liu, P. (2025). ‘Roles of arbuscular mycorrhizal fungi in plant growth and disease management for sustainable agriculture,’ Frontiers in Microbiology, 16, 1616273. Available at: https://doi.org/10.3389/fmicb.2025.1616273
______________________________________________
About the Authors
Adrián González Ortega-Villaizán
Adrián is a postdoctoral researcher at the Center for Plant Biotechnology and Genomics (CBGP) in Madrid (Spain), soon be joining the University of Nottingham (UK), and a 2026 Plantae Fellow . Adrián is a postdoctoral researcher at the Center for Plant Biotechnology and Genomics (CBGP) in Madrid (Spain) and he will soon be joining the University of Nottingham (UK). Find him on X: @adrigov98 | Bluesky: @adrigov.bsky.social.
Trevor Melusen
Trevor is currently a researcher at Plasmidsaurus and a 2026 Plantae Fellow. He focuses on making Next-Generation Sequencing faster and more affordable for all researchers. . You can find him on X: @trevor_melusen

