Silver birch: genome sequence and population genomic analyses
Plant Science Research Weekly, Research0 Comments
/
Salojärvi et al. have assembled the genome of silver birch (Betula pendula) and examined genomic diversity across 150 individuals collected from across Northern Europe and Asia. Their study shows that there have been several genetic bottlenecks associated with climate upheaval (e.g., at the K-Pg boundary…

Just add water: Could resurrection plants help feed the world?
GPC Blog, ResearchThis week we spoke to Professor Henk Hilhorst (Wageningen University and Research) about his research on desiccation tolerance in seeds and plants.
Could you begin by telling us a little about your research?
I am a plant physiologist specializing in seed biology. I have a long research…

A chromosome conformation capture ordered sequence of the barley genome
Plant Science Research Weekly, ResearchCereal grasses are of course economically important, but they also have large repetitive genomes with large pericentromeric regions that have been difficult to map and sequence. Barley (Hordeum vulgare L.) is used for human and animal food and fermented to produce beer and whisky. A barley sequence assembly…
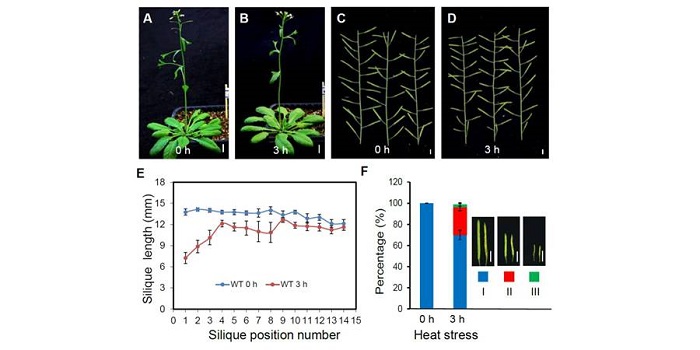
Tissue-specific transcriptomics shows the unfolded protein response’s role in maintaining fertility upon heat stress ($)
Plant Science Research Weekly, ResearchPlant reproduction is particularly sensitive to heat stress, so rising temperature is a major threat to food security. Zhang et al. surveyed the transcriptional responses to heat stress (3 h at 37º) in Arabidopsis and identified large differences between vegetative and reproductive tissue responses…
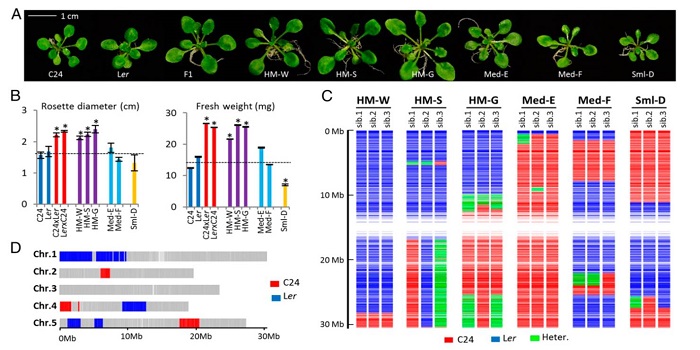
PIF4-controlled auxin pathway contributes to hybrid vigor in Arabidopsis thaliana
Plant Science Research Weekly, ResearchHybrid vigor is a well-known but still poorly understood phenomenon in which the F1 hybrid progeny of a cross often show enhanced growth as compared to either parent. True-breeding lines that retain this enhanced growth, known as hybrid mimics, have been developed and are important tools for understanding…

H2A monoubiquitination in Arabidopsis is generally independent of LHP1 and PRC2 activity
Plant Science Research Weekly, ResearchGene silencing by chromatin marks occurs in plants and animals, but there are often some differences in the details. Polycomb repressive complex 1 (PRC1) and PRC2 were first characterized in animals and shown to repress gene expression in part through histone modification; PRC1 has histone H2A E3 ubiquitin…
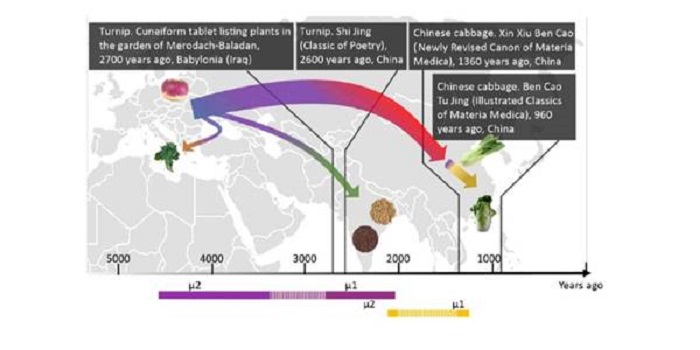
Genomic inferences of domestication events are corroborated by written records in Brassica rapa ($)
Plant Science Research Weekly, ResearchThere are many subspecies of Brassica rapa [including turnip (subsp. rapa) pak choi (subsp. chinensis) and Chinese cabbage (subsp. pekinensis)] but the relationships between the subspecies has remained uncertain. Qi et al. sequenced 143 accessions, including some subspecies for the first time. They then…
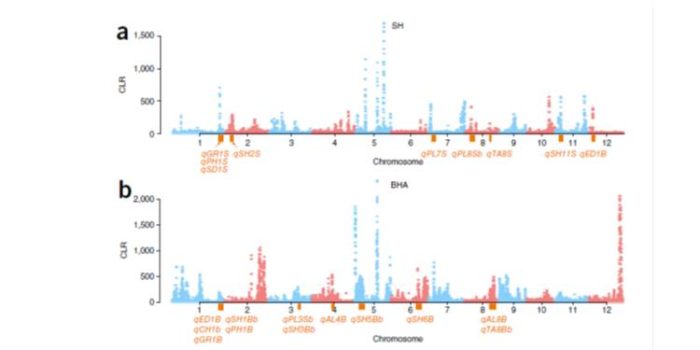
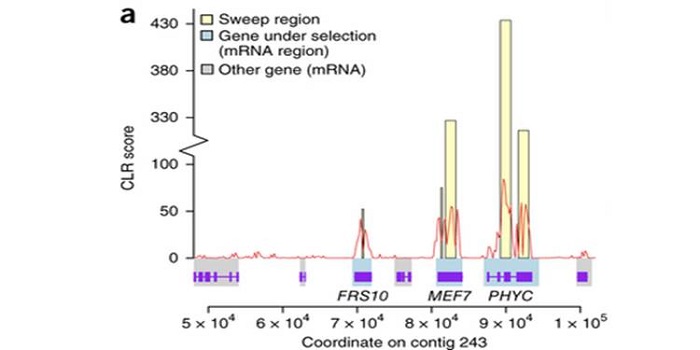
Signatures of adaptation in the weedy rice genome
Plant Science Research Weekly, ResearchCrop domestication has been accompanied by the evolution of aggressive weedy crop relatives that compete for resources and make weed management a challenge. By using whole-genome sequencing of the two most commonly found weedy rice strains in the US (SH and BHA), and comparing them with the genomes of…

Review: Insights into plant adaptation from transcriptomics and proteomics studies
Plant Science Research Weekly, ResearchOmics approaches have helped shed light on how plants relate to their environment and how they respond to changes in it. Although still relatively underutilized, comparative transcriptomics and proteomics approaches also can be applied to study mechanisms of plant adaptation. Voelckel et al. discuss…