OsKNAT7 bridges secondary cell wall formation and cell growth regulation
The plant cell wall is a complex structure composed of cellulose embedded in a matrix of polysaccharides (hemicellulose and pectin), phenolic compounds and proteins. The primary cell wall is a thin layer formed in growing cells and is present ubiquitously, whereas the secondary cell wall is a rigid and thick structure formed after cell growth in specialized cell types such as xylem vessels, fiber cells and anther endothecium (Zhong and Ye, 2014). Secondary cell walls originated when vascular plants appeared on land around 430 million years ago (Zhong et al., 2010). They provide rigidity to the upright plant body, enable water and nutrient transport through vascular tissues, and protect cells from microbial infection. The main components of the secondary cell wall are cellulose, hemicellulose (xylan and glucomannan) and lignin (Zhong and Ye, 2014). Secondary wall biosynthesis is regulated by an orchestra of transcription factors (TF), including NAC (NAM, NO APICAL MERISTEM; ATAF, ARABIDOPSIS TRANSCRIPTION ACTIVATION FACTOR; CUC, CUP-SHAPED COTYLEDON), MYB, KNOX (KNOTTED1-like homeobox) and BEL1-LIKE TFs (Rao and Dixon, 2018). In this issue of Plant Physiology, Wang et al. characterized the rice (Oryza sativa) class II KNOX TF KNOX ARABIDOPSIS THALIANA 7 (KNAT7) that coordinates secondary wall thickening and cell expansion in stem and rice grains.
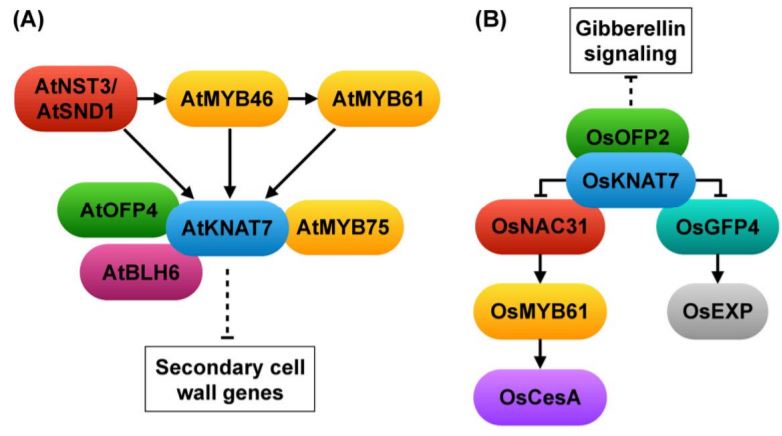
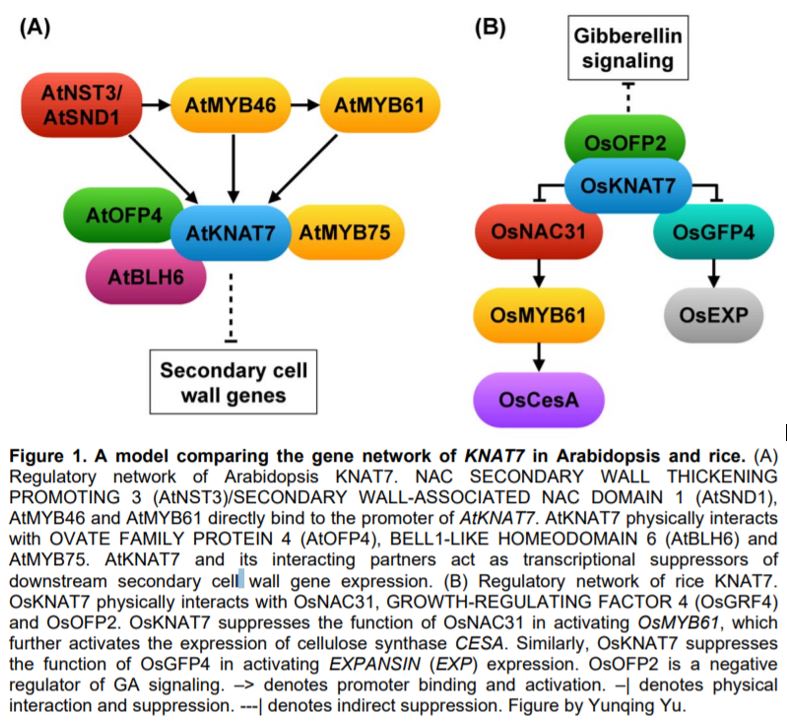 KNAT7 was first designated as IRREGULAR XYLEM11 based on its role in xylem development and secondary wall regulation in Arabidopsis thaliana (Brown et al., 2005). The knat7 knock-out mutant has thicker secondary cell walls and increased lignin content in fiber cells (Li et al., 2012). KNAT7 is a direct target of the secondary cell wall master regulator NAC SECONDARY WALL THICKENING PROMOTING 3 (NST3)/SECONDARY WALL-ASSOCIATED NAC DOMAIN 1 (SND1), MYB46 and MYB61 (Rao and Dixon, 2018). KNAT7 physically interacts with a number of transcription factors including OVATE FAMILY PROTEIN 4 (OFP4), BELL1-LIKE HOMEODOMAIN 6 (BLH6) and MYB75, and the protein complexes suppress the expression of secondary wall biosynthetic genes (Li et al., 2012; Nakano et al., 2015) (Fig. 1A).
KNAT7 was first designated as IRREGULAR XYLEM11 based on its role in xylem development and secondary wall regulation in Arabidopsis thaliana (Brown et al., 2005). The knat7 knock-out mutant has thicker secondary cell walls and increased lignin content in fiber cells (Li et al., 2012). KNAT7 is a direct target of the secondary cell wall master regulator NAC SECONDARY WALL THICKENING PROMOTING 3 (NST3)/SECONDARY WALL-ASSOCIATED NAC DOMAIN 1 (SND1), MYB46 and MYB61 (Rao and Dixon, 2018). KNAT7 physically interacts with a number of transcription factors including OVATE FAMILY PROTEIN 4 (OFP4), BELL1-LIKE HOMEODOMAIN 6 (BLH6) and MYB75, and the protein complexes suppress the expression of secondary wall biosynthetic genes (Li et al., 2012; Nakano et al., 2015) (Fig. 1A).
In this issue, Wang et al. showed that the KNAT7 ortholog in rice has a conserved function in regulating secondary cell wall development, but may accomplish this through different or additional interacting partners and downstream factors. The Osknat7 mutant has a thicker wall in fiber cells than the wild type, similar to the Arabidopsis knat7 mutant. However, the increased secondary wall is due to a higher content of cellulose and xylan, rather than lignin as in Arabidopsis (Li et al., 2012). The Osknat7 mutant also exhibits a larger grain size and larger cells in spikelet bracts (Wang et al., 2019), which has not been reported in Arabidopsis. The phenotype in both cell wall thickness and cell size suggests that OsKNAT7 may be a coordinator of secondary cell wall biosynthesis and cell expansion. Indeed, Wang et al. showed that OsKNAT7 physically interacts with both the secondary wall regulator NAC31, and the cell expansion regulator GROWTH-REGULATING FACTOR 4 (GRF4). Similar to the transcriptional repressor activity of AtKNAT7, OsKNAT7 suppresses the function of NAC31 and GRF4, which otherwise activate MYB61 and expansin gene expression, respectively (Wang et al., 2019) (Fig. 1B). In addition, OsKNAT7 interacts with OsOFP2 in a yeast two-hybrid assay; the latter regulates both grain shape and vascular bundle development via gibberellin signaling (Schmitz et al., 2015). The OsKNAT7-OsOFP2 complex could be another mechanism coordinating secondary wall thickening and cell growth (Fig. 1B).
In general, the transcriptional regulation of secondary cell wall biosynthesis is conserved in evolution. Numerous studies have shown that the ectopic expression of NAC or MYB master regulators from poplar (Populus trichocarpa), grasses, or gymnosperms is able to rescue the phenotype in mutants of Arabidopsis orthologues (Zhong et al., 2010; Rao and Dixon, 2018). However, the history of duplication in these TF families enables subfunctionalization and neofunctionalization, which may result in diverse regulatory mechanisms and cell wall compositions in different plant species and families. Indeed, the cell wall of the grass species is different from that of many dicots in that the grass cell wall has an increased amount of xylan, and that lignin and other cell wall components are often cross-linked with ferulic acid and p-coumaric acid (Hatfield et al., 2017). The finding in OsKNAT7 provides evidence that the gene network of KNAT7 may differ between Arabidopsis and rice. For instance, in Arabidopsis, MYB61 directly binds to the promoter of KNAT7 and thus acts upstream of KNAT7 (Fig. 1A), while in rice, KNAT7 suppresses NAC31 activation of MYB61 expression (Fig. 1B), suggesting that the order of signal transduction has changed during evolution. Alternatively, there could be a feedback loop between KNAT7 and MYB61 in both species. Further mechanistic studies will uncover the evolution of the KNAT7 network in balancing secondary cell wall formation with cell division and expansion.
Yunqing Yu, Assistant Features Editor
Donald Danforth Plant Science Center, Saint Louis, MO, USA
References
Brown DM, Zeef LA, Ellis J, Goodacre R, Turner SR (2005) Identification of novel genes in Arabidopsis involved in secondary cell wall formation using expression profiling and reverse genetics. The Plant Cell 17: 2281-2295
Hatfield RD, Rancour DM, Marita JM (2017) Grass cell walls: a story of cross-linking. Frontiers in plant science 7: 2056
Li E, Bhargava A, Qiang W, Friedmann MC, Forneris N, Savidge RA, Johnson LA, Mansfield SD, Ellis BE, Douglas CJ (2012) The Class II KNOX gene KNAT7 negatively regulates secondary wall formation in Arabidopsis and is functionally conserved in Populus. New Phytologist 194: 102-115
Nakano Y, Yamaguchi M, Endo H, Rejab NA, Ohtani M (2015) NAC-MYB-based transcriptional regulation of secondary cell wall biosynthesis in land plants. Frontiers in Plant Science 6: 288
Rao X, Dixon RA (2018) Current models for transcriptional regulation of secondary cell wall biosynthesis in grasses. Frontiers in Plant Science 9: 399
Schmitz AJ, Begcy K, Sarath G, Walia H (2015) Rice Ovate Family Protein 2 (OFP2) alters hormonal homeostasis and vasculature development. Plant Science 241: 177-188
Wang S, Yang H, Mei J, Liu X, Wen Z, Zhang L, Xu Z, Zhang B, Zhou Y (2019) A Rice homeobox protein KNAT7 integrates the pathways regulating cell expansion and wall stiffness. Plant Physiology: pp. 00639.02019
Zhong R, Lee C, Ye Z-H (2010) Evolutionary conservation of the transcriptional network regulating secondary cell wall biosynthesis. Trends in Plant Science 15: 625-632
Zhong R, Ye Z-H (2014) Secondary cell walls: biosynthesis, patterned deposition and transcriptional regulation. Plant and Cell Physiology 56: 195-214