Liguleless1, a conserved gene regulating leaf angle and a target for yield improvement in wheat
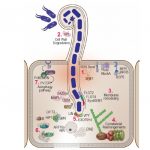
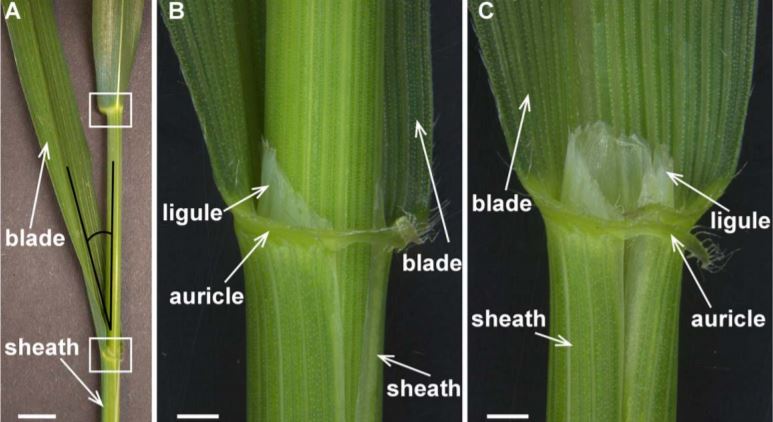
A major challenge in modern agriculture is to increase crop yield with diminishing agricultural land to feed the fast-growing global population. The erect leaf phenotype is a highly desirable trait that allows for higher planting density and maximizes light interception capacity and thus increases photosynthetic efficiency and crop yield (Donald, 1968). In cereal crops, which all belong to the grass family (Poaceae), a leaf is composed of a sheath, blade, and the junction between the two, called the laminar joint. The laminar joint includes the auricle, an appendage or expanded region at the base of the blade surrounding the sheath, and the ligule, which is the membranous outgrowth of the sheath (Fig. 1). Leaf angle is defined as the angle between the leaf blade midrib and the stem (Fig. 1A), and is influenced by leaf development, particularly at the laminar joint. Classic liguleless (lg) mutants, such as lg1, lg2, lg3 and lg4 in maize (Zea mays) and Oslg1 in rice (Oryza sativa), are deficient in the formation of the ligule and auricle, resulting in smaller leaf angles (Mantilla-Perez and Salas Fernandez, 2017). In this issue of Plant Physiology, Liu et al. show that the LG1 homolog in wheat (Triticum aestivum), TaSPL8, plays a conserved role in regulating leaf angle. They further demonstrate that TaSPL8 functions through regulating brassinosteroid biosynthesis and auxin signaling.
 Liu et al. (2019) reported a compact plant architecture wheat mutant with smaller laminar joint and vertical leaves identified from a fast neutron radiation mutagenesis screen. Map-based cloning combined with bulked segregant RNA sequencing narrowed down the candidates to 83 genes, among which TaSPL8, encoding a SQUAMOSA promoter-binding protein-like (SPL) transcription factor and homolog of LG1, was identified as the causal gene. Because wheat is a hexaploid, the authors generated CRISPR mutants to knock out all three TaSPL8 homoeologs. The homoeologs have additive effects on the phenotype, and the triple mutants completely lack the lamina joint. In addition, the contribution of the three homoeologs differs, with TaSPL8 from the D subgenome showing the largest effect (Liu et al., 2019). Overall, the phenotypes of Taspl8, particularly the higher order mutants, resemble the defects in the ligule and auricle development seen in maize and rice lg1 mutants (Becraft et al., 1990; Lee et al., 2007), suggesting that the functions of LG1/SPL8 are conserved across different grass species.
Liu et al. (2019) reported a compact plant architecture wheat mutant with smaller laminar joint and vertical leaves identified from a fast neutron radiation mutagenesis screen. Map-based cloning combined with bulked segregant RNA sequencing narrowed down the candidates to 83 genes, among which TaSPL8, encoding a SQUAMOSA promoter-binding protein-like (SPL) transcription factor and homolog of LG1, was identified as the causal gene. Because wheat is a hexaploid, the authors generated CRISPR mutants to knock out all three TaSPL8 homoeologs. The homoeologs have additive effects on the phenotype, and the triple mutants completely lack the lamina joint. In addition, the contribution of the three homoeologs differs, with TaSPL8 from the D subgenome showing the largest effect (Liu et al., 2019). Overall, the phenotypes of Taspl8, particularly the higher order mutants, resemble the defects in the ligule and auricle development seen in maize and rice lg1 mutants (Becraft et al., 1990; Lee et al., 2007), suggesting that the functions of LG1/SPL8 are conserved across different grass species.
ZmLG1 is specifically expressed at the preligule band, a linear layer of small cells at the boundary between the preblade and presheath. ZmLG1 activates the expression of auxin transporter ZmPIN1a, which induces auxin accumulation at the boundary and promotes ligule formation (Johnston et al., 2014). Leaf angle is regulated by brassinosteroid signaling, and exogenous 24-epibrassinolide promotes cell division and elongation of the adaxial cells at the laminar joint, resulting in increased leaf angles (Luo et al., 2016; Mantilla-Perez and Salas Fernandez, 2017). However, the link between LG1 and brassinosteroid signaling and direct targets of LG1 is largely unknown. Liu et al. (2019) screened for TaSPL8 targets in the down-regulated genes in the Taspl8 triple mutant compared to the wild-type, using the conserved SPL binding motif GTAC (Birkenbihl et al., 2005). Two candidates were identified: an AUXIN RESPONSE FACTOR 6 and a brassinosteroid biosynthetic gene D2. Direct binding of TaSPL8 to their promoters containing the GTAC motif was further validated by an electrophoretic mobility shift assay, chromatin immunoprecipitation followed by quantitative PCR in wheat, and a transient luciferase reporter assay in tobacco (Nicotiana tabacum) leaves. These results provide insight into the molecular function of LG1 in boundary establishment and laminar joint development.
The optimal leaf angle for maximal yield is unknown. A maize leaf blade has an average of leaf angle of 35° from the stem. Zmlg2, with a 14° leaf angle, has a significant increase in grain yield, particularly at higher planting density, while Zmlg1, with a more extreme leaf angle of 2°, does not have improved yield, suggesting that intermediate leaf angles may be more favorable than completely vertical leaves in crop breeding (Lambert and Johnson, 1978; Mantilla-Perez and Salas Fernandez, 2017). Although there is no difference in grain number per spike and grain weight between the wild-type and Taspl8 triple mutants, spike number (i.e., tiller number) is significantly higher in the triple mutants than in the wild-type, particularly at higher planting density, which may enhance yield. Future studies of grain yield in mutants with intermediate leaf angles may further benefit wheat breeding.
Yunqing Yu
Donald Danforth Plant Science Center
Saint Louis, MO, USA
References:
Becraft PW, Bongard-Pierce DK, Sylvester AW, Poethig RS, Freeling M (1990) The liguleless-1 gene acts tissue specifically in maize leaf development. Developmental Biology 141: 220-232
Birkenbihl RP, Jach G, Saedler H, Huijser P (2005) Functional dissection of the plant-specific SBP-domain: overlap of the DNA-binding and nuclear localization domains. Journal of Molecular Biology 352: 585-596
Donald CM (1968) The breeding of crop ideotypes. Euphytica 17: 385-403
Johnston R, Wang M, Sun Q, Sylvester AW, Hake S, Scanlon MJ (2014) Transcriptomic analyses indicate that maize ligule development recapitulates gene expression patterns that occur during lateral organ initiation. The Plant Cell 26: 4718-4732
Lambert R, Johnson R (1978) Leaf angle, tassel morphology, and the performance of maize hybrids 1. Crop Science 18: 499-502
Lee J, Park J-J, Kim SL, Yim J, An G (2007) Mutations in the rice liguleless gene result in a complete loss of the auricle, ligule, and laminar joint. Plant Molecular Biology 65: 487-499
Liu K, Cao J, Yu K, Liu X, Gao Y, Chen Q, Zhang W, Peng H, Du J, Xin M (2019) Wheat TaSPL8 Modulates Leaf Angle Through Auxin and Brassinosteroid Signaling. Plant Physiology: pp. 00248.02019
Luo X, Zheng J, Huang R, Huang Y, Wang H, Jiang L, Fang X (2016) Phytohormones signaling and crosstalk regulating leaf angle in rice. Plant cell reports 35: 2423-2433
Mantilla-Perez MB, Salas Fernandez MG (2017) Differential manipulation of leaf angle throughout the canopy: current status and prospects. Journal of Experimental Botany 68: 5699-5717