Insights into the Evolution of C4 Photosynthesis in the Grasses
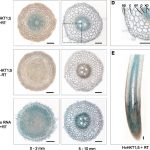
Most evidence supports the idea that the C4 photosynthetic pathway arose via a series of innovations that initially enabled plants to refix a large 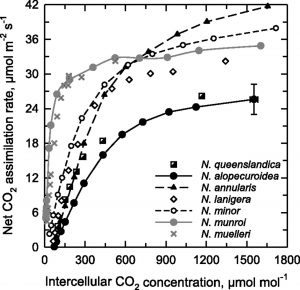 fraction of photorespired CO2 via the shuttling of Gly from the mesophyll (M) to a sheath of cells surrounding the vascular tissue, where the Gly is decarboxylated. The sheath layer is most commonly the bundle sheath (BS) cells. Associated with the shuttling of Gly is a shift in the expression of the mitochondrial enzyme Gly decarboxylase (GDC) from M cells to the vascular sheath tissue. This also requires that the Gly molecules produced in photorespiration diffuse from M cells to the sheath cells for metabolism. Gly shuttling, also known as C2 photosynthesis, elevates CO2 levels in the BS by a factor of two to three times relative to M cells, thereby improving Rubisco efficiency. The C2 condition is present in more than 40 intermediate species that branch between C3 and C4 species on the phylogenetic trees of more than a dozen distinct C4 lineages. The Australian grass subtribe Neurachninae is interesting to study because it contains closely related species that use C3, C4, and C2 photosynthesis. Since most previous research in this area has examined eudicot species, Khoshravesh et al. (10.1104/pp.19.00925) examined leaf gas exchange, anatomy and ultrastructure, and tissue localization of Gly decarboxylase subunit P (GLDP) in nine Neurachninae species. They identify previously unrecognized variations in leaf structure and physiology within the Neurachne that represents varying degrees of C3–C4 intermediacy. Their results are consistent with a model wherein C4 evolution in the Neurachninae initially occurred via an increase in organelle and GLDP content in MS cells, which generated a sink for photorespired CO2 in MS tissues. This research indicates the potential of the Neurachninae to provide novel insights into C4 evolution in grasses, particularly in the initial transition from a C3 character state.
fraction of photorespired CO2 via the shuttling of Gly from the mesophyll (M) to a sheath of cells surrounding the vascular tissue, where the Gly is decarboxylated. The sheath layer is most commonly the bundle sheath (BS) cells. Associated with the shuttling of Gly is a shift in the expression of the mitochondrial enzyme Gly decarboxylase (GDC) from M cells to the vascular sheath tissue. This also requires that the Gly molecules produced in photorespiration diffuse from M cells to the sheath cells for metabolism. Gly shuttling, also known as C2 photosynthesis, elevates CO2 levels in the BS by a factor of two to three times relative to M cells, thereby improving Rubisco efficiency. The C2 condition is present in more than 40 intermediate species that branch between C3 and C4 species on the phylogenetic trees of more than a dozen distinct C4 lineages. The Australian grass subtribe Neurachninae is interesting to study because it contains closely related species that use C3, C4, and C2 photosynthesis. Since most previous research in this area has examined eudicot species, Khoshravesh et al. (10.1104/pp.19.00925) examined leaf gas exchange, anatomy and ultrastructure, and tissue localization of Gly decarboxylase subunit P (GLDP) in nine Neurachninae species. They identify previously unrecognized variations in leaf structure and physiology within the Neurachne that represents varying degrees of C3–C4 intermediacy. Their results are consistent with a model wherein C4 evolution in the Neurachninae initially occurred via an increase in organelle and GLDP content in MS cells, which generated a sink for photorespired CO2 in MS tissues. This research indicates the potential of the Neurachninae to provide novel insights into C4 evolution in grasses, particularly in the initial transition from a C3 character state.