A Conserved Mechanism to Terminate Floral Meristems
In plants, the balance between stem cell renewal and loss is carefully maintained by intricate regulatory networks. Although the root and shoot apical meristems can in principle continue to grow and self-renew indefinitely, the floral meristem is terminated once flower formation is completed. The precise timing of floral meristem termination can have dramatic effects on the final form of the flower and fruit. Bollier et al. (2018) compare the mechanisms that govern floral meristem termination in two distantly related species, Arabidopsis and tomato.
It has been known for some time that, in Arabidopsis, the floral homeotic protein AGAMOUS (AG) controls the formation of stamens and carpels as well as floral meristem termination. The development of these reproductive organs is disturbed, or lost, in loss-of-function mutants of AG and its tomato homolog TOMATO AGAMOUS 1 (TAG1) while the flowers also produce reiterations of organs of the perianth (Bowman et al., 1989; Pneuli et al., 1994). In Arabidopsis, AG acts in meristem termination by suppressing the levels of WUSCHEL (WUS) through two compatible mechanisms. At early stages of flower development, AG recruits polycomb group proteins to the WUS locus to initiate its downregulation, although the activity of these proteins is not sufficient to completely suppress WUS expression (Liu et al., 2011). In Arabidopsis, AG also promotes the expression of another transcription factor, KNUCKLES (KNU), which also inhibits WUS expression (Sun et al., 2009). But questions remained about the conservation of KNU function in tomato and its interactions with other factors.
Bollier et al. (2018) addressed these questions by analyzing KNU activity in Arabidopsis and tomato. They showed that KNU function is conserved in tomato: KNU expression is promoted by TAG1 and plants with lower KNU activity bore fruits that contained up to 5 additional locules, indicating disruption of meristem termination (see figure). The authors also studied the Arabidopsis ortholog of another gene that was previously shown to regulate floral meristem termination in tomato (Sicard et al., 2008). They show that, similar to the corresponding mutant in tomato (Sicard et al., 2008), Arabidopsis plants lacking the activity of a MINI ZINC FINGER (MIF) family protein also lose determinacy. Through a series of biochemical assays, the authors show that KNU in both species interacts with the respective MIF protein and, as expected, WUS expression was elevated or persisted in plants lacking the activity of these proteins. In addition, several experiments provide evidence that KNU and MIF bind to the WUS locus in order to suppress its expression. The authors propose that the interaction between MIF and KNU recruits the transcriptional corepressor TOPLESS and HISTONE DEACETYLASE 19 to remove acetylation at the WUS locus.
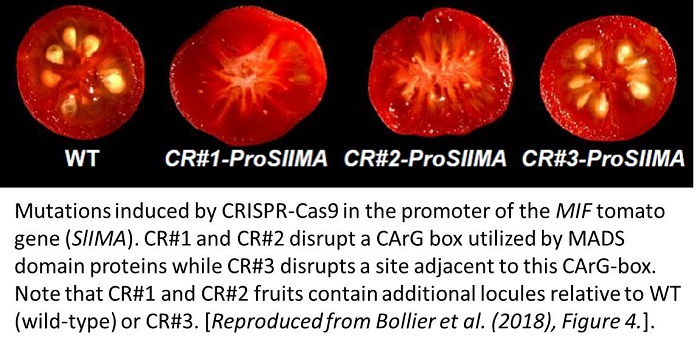
Conservation of this mechanism between Arabidopsis and tomato indicates that other angiosperms are likely to use a similar process to terminate floral meristems. This represents a clear target for genetic engineering: to disrupt binding sites for KNU or MIF in the WUS locus. Bollier et al. (2018) present an example of how the strategy of disrupting transcription factor binding sites can be effective by removing a MADS domain binding sequence in the promoter of the tomato MIF gene via CRISPR-Cas9 technology. Isolated mutants contained up to 6 locules rather than the maximum of 3 seen in wild-type counterparts. The identification of the MADS domain protein that interacts with this region, as well as furthering the understanding of how acetyl groups are removed from histones associated with WUS are key future challenges.
REFERENCES
Bollier N., Sicard A., Leblond J., Latrasse D., Gonzalez N., Gévaudant F., Benhame M., Raynaud C., Lenhard M., Chevalier C., Hernould M., and Delmas F. (2017) At-MINI ZINC FINGER2 and Sl-INHIBITOR OF MERISTEM ACTIVITY, a Conserved Missing Link in the Regulation of Floral Meristem Termination in Arabidopsis and Tomato. Plant Cell http://www.plantcell.org/content/early/2018/01/03/tpc.17.00653
Bowman J.L., Smyth D.R. and Meyerowitz E.M. (1989) Genes directing flower development in Arabidopsis. Plant Cell 1(1):37-52. https://doi.org/10.1105/tpc.1.1.37
Liu X., Xu Y., Müller R., Yumul R.E., Liu C., Pan Y., Cao X., Goodrich J. and Chen X. (2011) AGAMOUS Terminates Floral Stem Cell Maintenance in Arabidopsis by Directly Repressing WUSCHEL through Recruitment of Polycomb Group Proteins. Plant Cell (10):3654-70 http://www.plantcell.org/content/23/10/3654.long
Pnueli L., Hareven D., Rounsley S.D., Yanofsky M.F. and Lifschitz E. (1994) Isolation of the tomato AGAMOUS gene TAG1 and analysis of its homeotic role in transgenic plants. Plant Cell 6(2):163-73. https://doi.org/10.1105/tpc.6.2.163
Sicard A., Petit J., Mouras A., Chevalier C. and Hernould M. (2008) Meristem activity during flower and ovule development in tomato is controlled by the mini zinc finger gene INHIBITOR OF MERISTEM ACTIVITY. Plant J 55(3):415-27 http://onlinelibrary.wiley.com/doi/10.1111/j.1365-313X.2008.03520.x/abstract
Sun B., Xu Y., Ng K.H. and Ito T. (2009) A timing mechanism for stem cell maintenance and differentiation in the Arabidopsis floral meristem. Genes & Development 23(15):1791-804 http://genesdev.cshlp.org/content/23/15/1791.long








