APC/CTE Shapes Rice Architecture from Top to Bottom
Gibberellins (GAs) and abscisic acid (ABA) affect plant development in an opposite manner. GA is generally considered a growth promoting hormone, whereas ABA signaling, triggered in response to stress, counteracts the GA effects and restricts growth under suboptimal conditions (Vanstraelen and Benkova, 2012). To integrate the intrinsic and external cues, ABA and GA need to antagonize each other’s actions. For example, in Arabidopsis, ABA inhibits GA functions by reducing GA biosynthesis, which results in stabilization of DELLA proteins, the GA repressors (Zentella et al., 2007). In turn, it was shown in rice that GA antagonizes ABA signaling by enhancing the degradation of ABA receptors (Lin et al., 2015).
The GA-mediated inhibition of ABA signaling in rice requires an activating component of APC/CTE E3 ubiquitin ligase complex, TILLER ENHANCER (TE) (Lin et al., 2015). TE, by the physical interaction with ABA receptors OsPYL/RCARs, recruits them to APC/CTE complex, which subsequently targets them for proteasomal degradation. ABA activates SnRK2 protein kinases to phosphorylate TE, which releases OsPYL/RCARs from the interaction with TE and thus prevents their degradation. To antagonize ABA, increased GA levels reduce SnRK2 protein abundance, and as a result promote OsPYL/RCAR protein decay (Lin et al., 2015).
TE was initially identified as a negative regulator of tiller development (Lin et al, 2012). Recent work of Lin et al. (2020) demonstrates that APC/CTE is also a hub for ABA-GA antagonistic control of both root growth and shoot branching in rice. The authors showed that overexpression of TE resulted in a dramatic decrease in root length, indicating that APC/CTE regulates root growth. Lin et al. (2020) further investigated the involvement of ABA and GA in APC/CTE–mediated root growth. Low levels of GA stimulated root meristem (RM) cell proliferation and as a consequence enhanced root growth. By contrast, higher GA levels restricted RM cell division, resulting in root growth inhibition. Only low ABA doses enhanced root growth, but this enhancement was suppressed by elevated TE expression. In the shoot, GA repressed whereas ABA induced tillering, yet the latter was restrained by TE overexpression. Together, this shows that ABA and GA antagonism in root growth and shoot branching regulation is mediated by APC/CTE complex (Lin et al, 2020).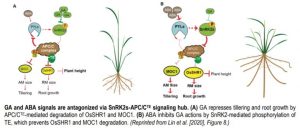
APC/CTE has previously been shown to mediate the decay of MOC1, a GRAS family transcription factor (TF) that induces tillering (Lin et al., 2012). Here, the authors screened sequences of known root growth regulators for a TE recognition motif and identified another GRAS TF, SHORTROOT1 (OsSHR1), as an APC/CTE substrate. Silencing OsSHR1 reduced RM cell division and as a consequence suppressed root growth, similarly to GA treatment (Lin et al., 2020).
APC/CTE -mediated degradation of OsSHR1 and MOC1 was induced by GA, but ABA was able to suppress this effect. GA enhanced physical interaction of TE with OsSHR1 and MOC1 and thus their recruitment to APC/CTE complex. Subsequent proteolysis of OsSHR1 and MOC1 diminished the activity of root and axillary meristems. In the presence of ABA, SnRK2 protein kinases phosphorylate TE, which decreases TE affinity to its substrates (Lin et al., 2015). By this mechanism, ABA prevents OsSHR1 and MOC1 from entering the APC/CTE complex, stabilizing their protein levels and thus meristematic activity.
Lin et al. (2020) present a model in which antagonistic action of ABA and GA integrates growth and development with external conditions via a SnRK2-APC/CTE signaling module (see figure). Interestingly, this SnRK2-APC/CTE hub mediates opposing effects of ABA and GA in both directions. GA can inhibit ABA actions via ABA receptor degradation (Lin et al., 2015), while ABA can antagonize GA signaling by stabilizing the levels of growth-related TFs (Lin et al., 2020). Both pathways rely on modulation of SnRK2-mediated phosphorylation of TE. Moreover, it presents the SnRK2-APC/CTE module as a critical mechanism regulating overall rice plant architecture, in both aerial and underground organs, as well as a putative dial for fine-tuning hormone-mediated growth.
Dorota Kawa
Department of Plant Biology and Genome Center
University of California, Davis
ORCID: 0000-0002-4227-1621
REFERENCES
Lin, Q., Wu, F., Sheng, P., Zhang, Z., Zhang, X., Guo, X., Wang, J., Cheng, Z., Wang, J., Wang, H., and Wan, J. (2015). The SnRK2-APC/C(TE) regulatory module mediates the antagonistic action of gibberellic acid and abscisic acid pathways. Nat Commun 6, 7981.
Lin, Q., Wang, D., Dong, H., Gu, S., Cheng, Z., Gong, J., Qin, R., Jiang, L., Li, G., Wang, J.L., Wu, F., Guo, X., Zhang, X., Lei, C., Wang, H., and Wan, J. (2012). Rice APC/C(TE) controls tillering by mediating the degradation of MONOCULM 1. Nat Commun 3, 752.
Vanstraelen, M., and Benkova, E. (2012). Hormonal interactions in the regulation of plant development. Annu Rev Cell Dev Biol 28, 463-487.
Zentella, R., Zhang, Z.L., Park, M., Thomas, S.G., Endo, A., Murase, K., Fleet, C.M., Jikumaru, Y., Nambara, E., Kamiya, Y., and Sun, T.P. (2007). Global analysis of della direct targets in early gibberellin signaling in Arabidopsis. Plant Cell 19, 3037-3057.