An evolutionary perspective on LysM receptors reveals conserved mechanisms for microbial signal perception
Lena Maria Müller
Boyce Thompson Institute
[email protected]
Plants are in constant contact with millions of microbes, which can be pathogenic or establish mutualistic interactions with the plant. One well-studied mutualistic interaction is the nodulation symbiosis of legumes with nitrogen-fixing rhizobia, which are harbored in specialized root organs called nodules (Parniske, 2018). In addition to legumes, several other plant species of the clades Fagales, Rosales, and Cucurbitales can form nodulation symbiosis predominantly with actinobacteria of the genus Frankia. Furthermore, most land plants can engage in mutually beneficial interactions with soil fungi in a symbiosis called arbuscular mycorrhiza (AM).
Plants perceive microbial signals by cell-surface receptors such as lysine-motif (LysM)-containing receptors. Such signals include chitin oligomers produced by pathogenic microbes, or lipo-chitooligosaccharides (LCOs) secreted by mutualistic rhizobia and AM fungi. LysM receptors belong to a large protein family that can be subdivided into multiple clades (Buendia et al., 2018). In legumes, rhizobial LCOs are perceived by a heteromeric complex consisting of LysM-type receptors of the LYK-I and LYR clades. Both receptor types are characterized by extracellular LysM domains; however, only LYK-I-type (but not LYR-type) receptors also contain a functional intracellular kinase domain. The Medicago truncatula LYK-I and LYR-type receptors involved in rhizobial LCO signaling are called LYSM DOMAIN CONTAINING RECEPTOR KINASE 3 (MtLYK3) and NOD FACTOR PERCEPTION (MtNFP), respectively (Moling et al., 2014). LCO perception by these receptors activates an intracellular signaling pathway culminating in nuclear calcium oscillations and ultimately leading to the expression of genes required for nodule formation (Oldroyd, 2013) (Fig. 1A). By contrast, chitin oligomers derived from pathogenic microbes are perceived by a protein complex containing CERK1-like LysM receptors and activate an immune response in the plant (Miya et al., 2007). Several LysM receptors, including MtCERK1, have dual functions in symbiosis and immunity signaling (Feng et al., 2019), which raises the question of how plants distinguish between symbionts and pathogens, and if chitin receptors evolved from LCO receptors or vice versa.
In this issue of Plant Physiology, Rutten et al. (2020) report an evolutionary perspective on LCO perception by studying LysM-type receptors in the tropical tree Parasponia andersonii (order Rosales). Parasponia is the only known non-legume that forms nodulation symbiosis with rhizobia. Parasponia and legumes diverged ~100 million years ago at the root of the nitrogen-fixing clade (van Velzen et al., 2018) and thus, Parasponia is ideally suited for comparative evolutionary analyses of symbiotic LysM receptors.
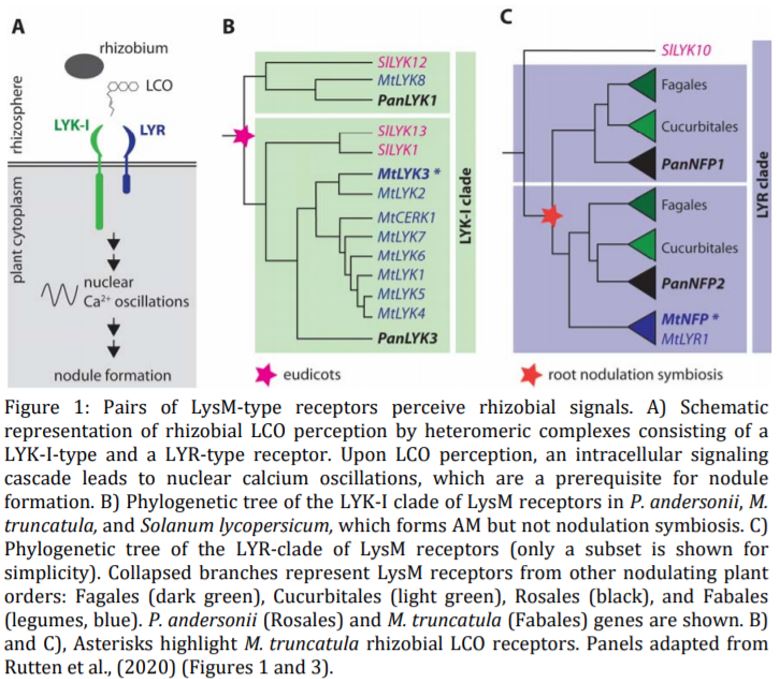 First, Rutten et al. (2020) examined the phylogeny of LYK-I-type receptors and detected evidence for an ancient duplication event, which occurred in an ancestral eudicot and gave rise to two distinct orthogroups within this clade of LysM receptors (Fig. 1B). The P. andersonii genome encodes one LYK-I-type receptor of each orthogroup, PanLYK1 and PanLYK3, respectively. PanLYK3 is the closest ortholog of the M. truncatula LCO receptor MtLYK3, but also of the immunity-associated chitin receptor MtCERK1. In addition, Rutten et al. (2020) identified two P. andersonii genes in the LYR clade, which they annotated as PanNFP1 and PanNFP2. PanNFP2 is the closest ortholog of MtNFP. PanNFP1 and PanNFP2 arose from an ancient duplication event coinciding with the origin of nodulation symbiosis (Fig. 1C).
First, Rutten et al. (2020) examined the phylogeny of LYK-I-type receptors and detected evidence for an ancient duplication event, which occurred in an ancestral eudicot and gave rise to two distinct orthogroups within this clade of LysM receptors (Fig. 1B). The P. andersonii genome encodes one LYK-I-type receptor of each orthogroup, PanLYK1 and PanLYK3, respectively. PanLYK3 is the closest ortholog of the M. truncatula LCO receptor MtLYK3, but also of the immunity-associated chitin receptor MtCERK1. In addition, Rutten et al. (2020) identified two P. andersonii genes in the LYR clade, which they annotated as PanNFP1 and PanNFP2. PanNFP2 is the closest ortholog of MtNFP. PanNFP1 and PanNFP2 arose from an ancient duplication event coinciding with the origin of nodulation symbiosis (Fig. 1C).
Next, Rutten et al. (2020) sought to determine the function of the P. andersonii LysM-type receptors in the context of root nodule symbiosis. By trans-complementation of a Lotus japonicus LCO-receptor mutant (Miwa et al., 2006), Rutten et al. (2020) could show that all four Parasponia LysM receptors were able to elicit nuclear calcium oscillations upon LCO treatment. To study the function of Parasponia LysM-type receptors in nodule development, Rutten et al. (2020) then created single and double receptor mutants using CRISPR-Cas9-mediated mutagenesis in P. andersonii. While Pannfp2 mutants were not able to form any nodules, Pannfp1 and Panlyk3 mutants displayed reduced nodulation efficiency. In addition, a Panlyk1,Panlyk3 double mutant did not nodulate in the presence of rhizobia. These results indicate that all four tested Parasponia LysM-type receptors function as LCO receptors and affect nodulation symbiosis with rhizobia, albeit to different degrees.
Similar to rhizobia, AM fungi also produce LCOs (Maillet et al., 2011). Initiation of AM symbiosis and nodulation relies on a common intracellular signaling machinery ultimately leading to nuclear calcium oscillations in the host root (Oldroyd, 2013). However, research in legumes revealed that AM fungal and rhizobial LCOs are perceived by at least partially independent LysM receptor complexes (Feng et al., 2019). To determine if such a functional diversification of LysM receptors also exists in Parasponia, Rutten et al. (2020) tested the response of their CRISPR mutants to AM fungi. While PanNFP1 and PanNFP2 were not found to be involved in AM symbiosis formation, the Panlyk1, Panlyk3 double mutant was severely impaired in establishing AM symbiosis, and the double mutants showed only few fungal infections. This indicates at least a partial functional overlap of the receptors responsible for the perception of rhizobial and AM fungal LCOs in P. andersonii. Interestingly, Panlyk3 mutants were also impaired in eliciting an immune response upon treatment with chitin heptamers, indicating that PanLYK3, in addition to its function in nodulation and AM symbiosis, is required for immune signaling.
Taken together, this research provides an interesting perspective on LysM receptor evolution. Rutten et al. (2020) report that legumes and Parasponia employ orthologous LYR- and LYK-I-type receptors to perceive rhizobial LCOs. While the Parasponia LYR-type receptor function was specific to nodulation symbiosis, Parasponia LYK-I receptors displayed a lower functional specificity, as—in addition to nodulation—they also mediate AM symbiosis (PanLYK1 and PanLYK3) and chitin perception (PanLYK3). Because PanLYK1 and PanLYK3 originated from an ancient duplication event that occurred prior to symbiotic nitrogen fixation, it is likely that the ancestral function of the LYK-I receptor clade is perception of LCOs, possibly those of AM fungi or other microbes. It is conceivable that the recent expansion of this receptor clade in legumes led to a functional separation of microbial signal perception (Fig. 1B). It will be interesting to decipher how Parasponia, which encodes fewer receptors than legumes with partially overlapping functions, differentiates between symbiosis and immunity signaling.
Literature Cited
Buendia L, Girardin A, Wang T, Cottret L, Lefebvre B (2018) LysM Receptor-Like Kinase and LysM Receptor-Like Protein Families: An Update on Phylogeny and Functional Characterization. Front Plant Sci 9: 539
Feng F, Sun J, Radhakrishnan GV, Lee T, Bozsoki Z, Fort S, Gavrin A, Gysel K, Thygesen MB, Andersen KR, et al (2019) A combination of chitooligosaccharide and lipochitooligosaccharide recognition promotes arbuscular mycorrhizal associations in Medicago truncatula. Nat Commun 10: 1–12
Maillet F, Poinsot V, André O, Puech-Pagès V, Haouy A, Gueunier M, Cromer L, Giraudet D, Formey D, Niebel A, et al (2011) Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 469: 58–63
Miwa H, Sun J, Oldroyd GED, Downie JA (2006) Analysis of Nod-Factor-Induced Calcium Signaling in Root Hairs of Symbiotically Defective Mutants of Lotus japonicus. Mol Plant Microbe Interact 19: 914–923
Miya A, Albert P, Shinya T, Desaki Y, Ichimura K, Shirasu K, Narusaka Y, Kawakami N, Kaku H, Shibuya N (2007) CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proceedings of the National Academy of Sciences 104: 19613–19618
Moling S, Pietraszewska-Bogiel A, Postma M, Fedorova E, Hink MA, Limpens E, Gadella TWJ, Bisseling T (2014) Nod Factor Receptors Form Heteromeric Complexes and Are Essential for Intracellular Infection in Medicago Nodules. The Plant Cell 26: 4188–4199
Oldroyd GED (2013) Speak, friend, and enter: signalling systems that promote beneficial symbiotic associations in plants. Nat Rev Microbiol 11: 252–263
Parniske M (2018) Uptake of bacteria into living plant cells, the unifying and distinct feature of the nitrogen-fixing root nodule symbiosis. Current Opinion in Plant Biology 44: 164–174
van Velzen R, Holmer R, Bu F, Rutten L, van Zeijl A, Liu W, Santuari L, Cao Q, Sharma T, Shen D, et al (2018) Comparative genomics of the nonlegume Parasponia reveals insights into evolution of nitrogen-fixing rhizobium symbioses. Proceedings of the National Academy of Sciences 115: E4700–E4709






