The Healing Power of Light
Light is life, especially for plants. It fuels photosynthesis and, when perceived by photoreceptors, directs important developmental programs, including photomorphogenesis and shade avoidance. Red and blue light wavelengths activate phytochrome (phy) and cryptochrome (cry) photoreceptors, respectively, which repress transcription factors that antagonize photomorphogenic development, such as the PHYTOCHROME INTERACTING FACTORs (PIFs). When phys and crys are inactive, in darkness or in vegetation shade, active PIFs induce stem elongation but repress leaf expansion and greening. Seasonal fluctuations in daylength also regulate plant growth, as the longer nights during winter time allow PIFs to accumulate (Gommers and Monte, 2018). In general, photoreceptor inactivity can be associated with stem elongation (Fig. 1).
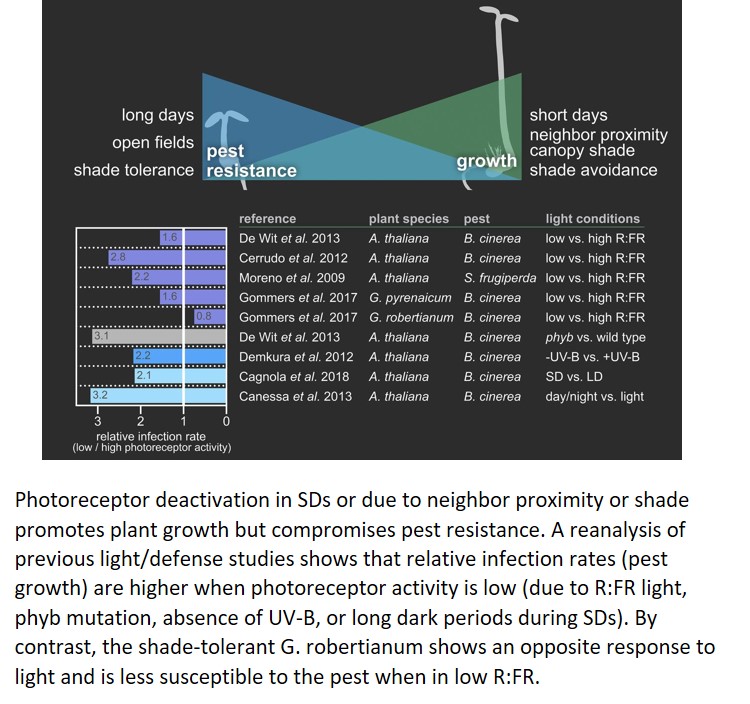
This elongation comes with a cost. For example, the shade avoidance syndrome (SAS; induced by neighboring plants or canopy shade) is characterized by strong stem growth but accompanied by increased susceptibility to several pests (Fig. 1; Moreno et al., 2009; Cerrudo et al., 2012; de Wit et al., 2013). Induced defenses mediated by the hormones salicylic acid and jasmonic acid (JA) are actively repressed when the plant strives for light in a canopy. It appears that there is a trade-off where the fight for light is prioritized over the fight against offenders. The mechanisms by which light perception affects JA-mediated pathways are very well described and extensively reviewed (Ballaré, 2014). In short, in sunlight (when the red to far-red [R:FR] light ratio is high), phys are active and indirectly stabilize DELLA proteins. These DELLA proteins bind to and inhibit JASMONATE-ZIM-DOMAIN (JAZ) proteins, inhibitors of JA-mediated defenses. When far-red light is enriched (low R:FR, due to reflection from neighboring leaves), phys are inactive and JAZ proteins are released, which makes plants more susceptible to, for example, the necrotrophic pathogen Botrytis cinerea or insect herbivores.
This growth defense trade-off is common among shade-intolerant species, but some shade-tolerant plants lack the SAS and even increase resistance against B. cinerea when far-red light is enriched (Fig. 1; Gommers et al., 2017). This response may help these species to thrive in the deep shade where avoiding shade is impossible and carbon gain is limited, so that conserving their energy resources is essential (Valladares and Niinemets, 2008).
A similar growth defense trade-off is seen when plants are exposed to UV-B light. UV-B, perceived by the UVR8 receptor, inhibits Arabidopsis (Arabidopsis thaliana) growth while making it more resistant to B. cinerea (Fig. 1). Unlike phy, UVR8-mediated induction of resistance seems to act independently of JA (Demkura and Ballaré, 2012).
Although the effect of light quality on plant susceptibility to pathogens and herbivores is quite well known, the influence of daylength has been less clear. In this edition of Plant Physiology, Juan I. Cagnola and colleagues show that daylength strongly influences resistance to pathogens (Cagnola et al., 2018). By performing transcriptome analyses using short day (SD)-grown Arabidopsis plants that were exposed to an extended day (long day [LD]) before sampling, the authors found many differentially regulated genes associated with JA responses. They also found increased resistance to B. cinerea in LD-grown plants compared with those grown in SD. This resistance was mediated by the photoreceptors crys and phyA, which stabilize DELLAs in a COP1-dependent manner and release JA-mediated defenses, similar to the pathways known from the SAS studies. Previously, by comparing plants grown in continuous light versus day/night rhythms, a similar correlation between hours of light and infection rate was found by Canessa et al. (2013); Fig. 1).
Summarizing these studies, we can conclude that in environments where, over a 24-h period, active photoreceptors are accumulating (e.g. LDs and open fields), stem elongation is repressed and plant defense pathways work optimally (Fig. 1). On the contrary, when inactive photoreceptors are abundant (e.g. SDs, close plant proximity, and shade), stem elongation is induced and resistance is repressed (Fig. 1). Despite the plethora of data, several questions remain unanswered. For example, how do high temperatures, which naturally occur simultaneously with LDs, open fields, and high UV-B light and are perceived by phys (Legris et al., 2016), affect plant resistance to pests? And what is the effect of all these factors on the activity of the pest? Some studies indicate that B. cinerea growth and virulence are affected by daylength (Canessa et al., 2013). Also, can we really speak of a trade-off, associated with selective energy allocation between growth and defense mechanisms? Finally, is the reverse trade-off as seen in Geranium robertianum common for all shade-tolerant plants? And does it also exist in plants that evolved to grow in specific daylengths?
In short, the new data of Cagnola et al. support the theory that light is beneficial for plant health. And honestly, don’t many of us feel much healthier in summer, when days are long, the sun warms our skin, and we’re not locked up inside? That is, as long as drinks are plentiful, but I’ll leave that topic to the drought stress experts.
REFERENCES
Ballaré CL (2014) Light regulation of plant defense. Annu Rev Plant Biol 65: 335–363
Cagnola JI, Cerdán PD, Pacín M, Andrade A, Rodriguez V, Zurbriggen MD, Legris M, Buchovsky S, Carrillo N, Chory J, et al. (2018) Long-day photoperiod enhances jasmonic acid-related plant defense. Plant Physiol 178: 163–173
Canessa P, Schumacher J, Hevia MA, Tudzynski P, Larrondo LF (2013) Assessing the effects of light on differentiation and virulence of the plant pathogen Botrytis cinerea: characterization of the White Collar Complex. PLoS One 8: e84223
Cerrudo I, Keller MM, Cargnel MD, Demkura PV, de Wit M, Patitucci MS, Pierik R, Pieterse CMJ, Ballaré CL (2012) Low red/far-red ratios reduce Arabidopsis resistance to Botrytis cinerea and jasmonate responses via a COI1-JAZ10-dependent, salicylic acid-independent mechanism. Plant Physiol 158: 2042–2052
Demkura PV, Ballaré CL (2012) UVR8 mediates UV-B-induced Arabidopsis defense responses against Botrytis cinerea by controlling sinapate accumulation. Mol Plant 5: 642–652
de Wit M, Spoel SH, Sanchez-Perez GF, Gommers CMM, Pieterse CMJ, Voesenek LACJ, Pierik R (2013) Perception of low red:far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences in Arabidopsis. Plant J 75: 90–103
Gommers CMM, Monte E (2018) Seedling establishment: a dimmer switch-regulated process between dark and light signaling. Plant Physiol 176: 1061–1074
Gommers CMM, Keuskamp DH, Buti S, van Veen H, Koevoets IT, Reinen E, Voesenek LACJ, Pierik R (2017) Molecular profiles of contrasting shade response strategies in wild plants: differential control of immunity and shoot elongation. Plant Cell 29: 331–344
Legris M, Klose C, Burgie ES, Rojas CCR, Neme M, Hiltbrunner A, Wigge PA, Schäfer E, Vierstra RD, Casal JJ (2016) Phytochrome B integrates light and temperature signals in Arabidopsis. Science354: 897–900
Moreno JE, Tao Y, Chory J, Ballaré CL (2009) Ecological modulation of plant defense via phytochrome control of jasmonate sensitivity. Proc Natl Acad Sci USA 106: 4935–4940
Valladares F, Niinemets Ü (2008) Shade tolerance, a key plant feature of complex nature and consequences. Annu Rev Ecol Evol Syst 39: 237–257