Found: Middle Men in the Hormone Joint Task Force
Bethany Huot, Michigan State University
Driving through the country on a lazy summer day with a nice gentle breeze, the lush fields of wheat, corn and soybean appear peaceful. What you cannot see is the quiet war plants are fighting against the insects and pathogens that have pulled up to a feast of fresh veggies. Unfortunately, crop plants are at a competitive disadvantage with their wild cousins, as breeding for traits like large, juicy tomatoes and plump grain has left them partially disarmed (Dwivedi et al., 2016). Global crop losses due to pests and pathogens are estimated at 10 – 40%, depending on a number of variables, and are most severe in regions where food insecurity is highest (Savary et al., 2019). By studying wild or weedy species, like Arabidopsis thaliana, scientists hope to identify more of the protective barriers and weapons plants have designed to defend themselves to re-arm our crops.
Pathogens use different modes of attack; therefore, plants must have more than one trick up their leaves. Using specific hormone cocktails, plants trigger defense signaling pathways tailored to meet the current threat (Pieterse et al., 2012, Li et al., 2019). For example, a nice blend of jasmonate (JA) and ethylene (ET) is used to counter necrotrophic pathogens that kill plant cells to devour the nutritious contents they release. Ever taken an aspirin? Then you have sampled salicylic acid (SA), the primary hormone used to defend against biotrophic pathogens that try to sneak in undetected to pilfer sweet treats from living cells. We typically think of SA and JA/ET pathways as being mutually antagonistic, but this is the two-finger piano version of what is more like a symphony. The signaling intricacies underlying these responses are still being explored, with transcription factors high on the “Most Wanted” list as they are the gatekeepers for downstream signaling to occur.
In this issue of Plant Physiology, Zheng et al. (2019) reveal a member of the defense team, BTB AND TAZ DOMAIN PROTEIN 4 (BT4), to have an important role in funneling signals from SA and ET to trigger defense against the hemi-biotrophic pathogen, Pseudomonas syringae pv. tomato DC3000 (Pst DC3000) (Zheng et al., 2019). Proteins in the BT family are middle men in the signaling cascade, being induced by Ca2+, H2O2 and SA and then passing the message on by helping activate the expression of downstream defense genes (Du & Poovaiah, 2004).
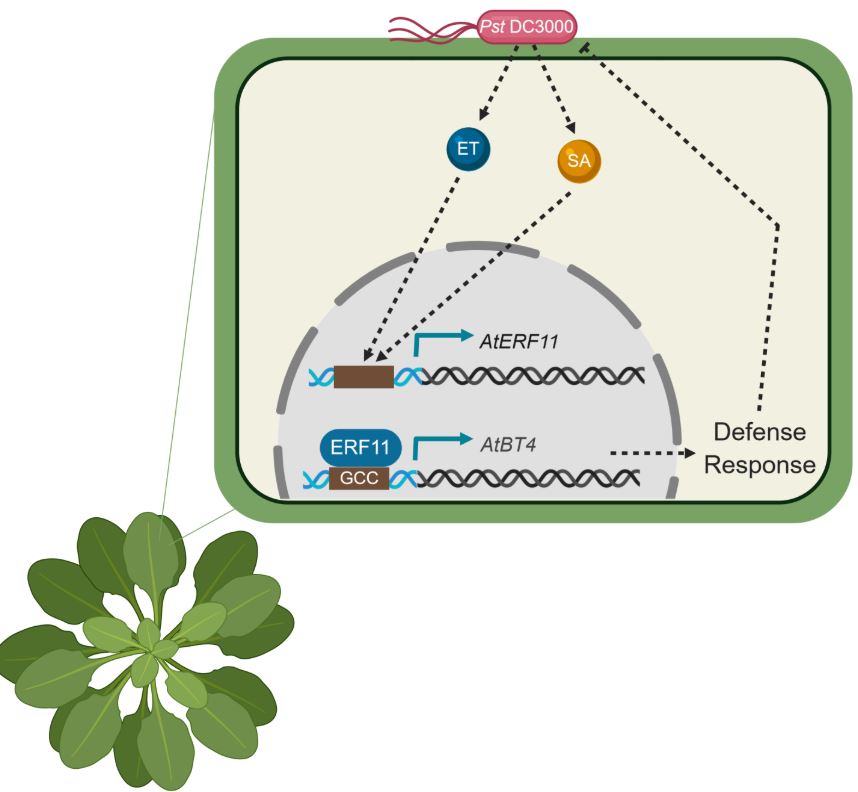
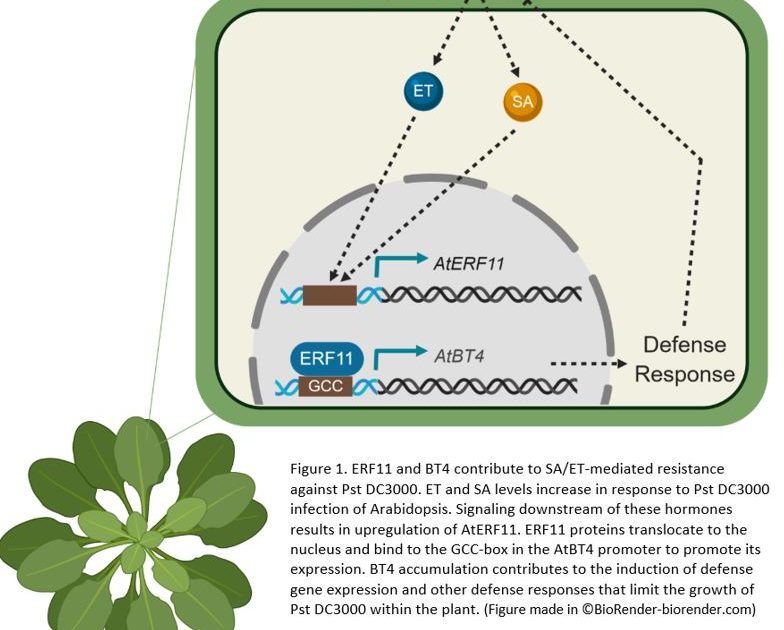 After establishing that both basal and SA-mediated protection against Pst DC3000 is compromised in bt4 mutant plants, the next mystery to be solved was how and where BT4 fits in the signaling pathway. Using a series of mutant plants missing key players important for SA and ET signaling, the research team show that BT4 expression is activated by both SA and ET in response to Pst DC3000 infection. How is this done? Well, our investigators hypothesized that one of the 122 ETHYLENE RESPONSE FACTOR (ERF) transcription factors might be the key. Indeed, their search turned up ERF11 as inducible by SA, ET and Pst DC3000. Further analyses showed that ERF11 is vital for activating BT4 in response to SA and ET via direct binding to the GCC box in the BT4 promoter (see Figure 1).
After establishing that both basal and SA-mediated protection against Pst DC3000 is compromised in bt4 mutant plants, the next mystery to be solved was how and where BT4 fits in the signaling pathway. Using a series of mutant plants missing key players important for SA and ET signaling, the research team show that BT4 expression is activated by both SA and ET in response to Pst DC3000 infection. How is this done? Well, our investigators hypothesized that one of the 122 ETHYLENE RESPONSE FACTOR (ERF) transcription factors might be the key. Indeed, their search turned up ERF11 as inducible by SA, ET and Pst DC3000. Further analyses showed that ERF11 is vital for activating BT4 in response to SA and ET via direct binding to the GCC box in the BT4 promoter (see Figure 1).
While ERFs are known to be induced under different stress conditions (Xie et al., 2019), Zheng et al. (2019) have connected the dots between ERF11, BT4 and classical defense signaling, shedding light on how ET and SA join forces to defend against a hemi-biotrophic pathogen. Plant scientists have spent decades working to decipher the elaborate code of hormone crosstalk in the hope of optimizing crop resilience under increasingly variable weather and soil conditions. Identifying these middle men in plant defense signaling brings us one step closer to achieving our goal so more of those delicious fruits, cereals and veggies will make it to our plates rather than being devoured by pesky insects and pathogens.
Figure 1. ERF11 and BT4 contribute to SA/ET-mediated resistance against Pst DC3000. ET and SA levels increase in response to Pst DC3000 infection of Arabidopsis. Signaling downstream of these hormones results in upregulation of AtERF11. ERF11 proteins translocate to the nucleus and bind to the GCC-box in the AtBT4 promoter to promote its expression. BT4 accumulation contributes to the induction of defense gene expression and other defense responses that limit the growth of Pst DC3000 within the plant. Figure made in ©BioRender – biorender.com
References
Du LQ, Poovaiah BW, 2004. A novel family of Ca2+/calmodulin-binding proteins involved in transcriptional regulation: interaction with fsh/Ring3 class transcription activators. Plant Molecular Biology 54, 549-69.
Dwivedi SL, Ceccarelli S, Blair MW, Upadhyaya HD, Are AK, Ortiz R, 2016. Landrace germplasm for improving yield and abiotic stress adaptation. Trends in Plant Science 21, 31-42.
Li N, Han X, Feng D, Yuan DY, Huang LJ, 2019. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: do we understand what they are whispering? International Journal of Molecular Sciences 20, 15.
Pieterse CMJ, Van Der Does D, Zamioudis C, Leon-Reyes A, Van Wees SCM, 2012. Hormonal modulation of plant immunity. Annual Review of Cell and Developmental Biology 28, 489-521.
Savary S, Willocquet L, Pethybridge SJ, Esker P, Mcroberts N, Nelson A, 2019. The global burden of pathogens and pests on major food crops. Nature Ecology & Evolution 3, 430-9.
Xie ZL, Nolan TM, Jiang H, Yin YH, 2019. AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis. Frontiers in Plant Science 10, 17.
Zheng X, Xing J, Zhang K, et al., 2019. Ethylene response factor ERF11 activates BT4 transcription to regulate immunity to Pseudomonas syringae. Plant Physiology, pp.01209.2018.