Photodamaged Chloroplasts Are Targets of Cellular Garbage Disposal
IN BRIEF by Gregory Bertoni [email protected]
 Autophagy, or “self eating,” is the process cells use to consume unwanted intracellular structures such as damaged organelles, excess membranes, and unneeded cellular proteins (Mizushima and Komatsu, 2011). Typically, the unwanted structure becomes surrounded by an autophagosomal membrane, which then fuses with the membrane of either the vacuole (yeast and plants) or the lysosome (animals) to deliver its contents for destruction by hydrolytic enzymes (Nakatogawa et al., 2009).
Autophagy, or “self eating,” is the process cells use to consume unwanted intracellular structures such as damaged organelles, excess membranes, and unneeded cellular proteins (Mizushima and Komatsu, 2011). Typically, the unwanted structure becomes surrounded by an autophagosomal membrane, which then fuses with the membrane of either the vacuole (yeast and plants) or the lysosome (animals) to deliver its contents for destruction by hydrolytic enzymes (Nakatogawa et al., 2009).
Mitochondria can be damaged by excess oxidation and thus may need to be destroyed. Mitophagy, the process by which cells consume defective mitochondria, is well characterized in both animals and yeast (Green et al., 2011; Youle and Narendra, 2011). Defects in mitophagy in humans can cause Parkinson’s-like symptoms, perhaps because impaired mitophagy results in accumulation of reactive oxygen species (ROS) and mutation of mitochondrial DNA.
During photosynthesis, excess natural or artificial light can cause photooxidative damage to chloroplasts. Izumi et al (2017) now show that chlorophagy, the intracellular destruction of chloroplasts, is induced by exposure to UVB irradiation (280 to 315 nm) and high light. They also demonstrate that entire photodamaged chloroplasts are transported to the plant cell vacuole for destruction. During leaf senescence and energy starvation, chloroplast stroma proteins are degraded by a type of autophagosome known as the Rubisco-containing body (Ishida et al., 2008). Whether a similar process operates in autophagy of photodamaged chloroplasts is a key focus of this work.
The authors first examine the relationship between UVB-induced damage and autophagy in WT vs. atg mutants, which are deficient in core proteins required for autophagy. Mutant plants irradiated with a UVB treatment as short as 1 h had less than 40% the shoot fresh weight than similarly treated wild-type plants, and they showed a similar reduction in photosynthetic performance. This demonstrates that Arabidopsis plants are more damaged by UVB when the autophagy process is defective.
Chloroplast location was monitored via expression of a chloroplast stroma-targeted GFP fusion protein (CT-GFP). Chloroplasts in untreated control leaves contained CT-GFP, but UVB exposure caused the appearance of CT-GFP-deficient chloroplasts in the central area of mesophyll cells where the vacuole is typically found. Simultaneous expression of a vacuolar membrane marker protein and a stromal RFP fusion protein showed that RFP-deficient chloroplasts were indeed found inside the vacuole. This was not the case in atg mutant plants, showing that their movement to the vacuole depends on the presence of a functional autophagy pathway.
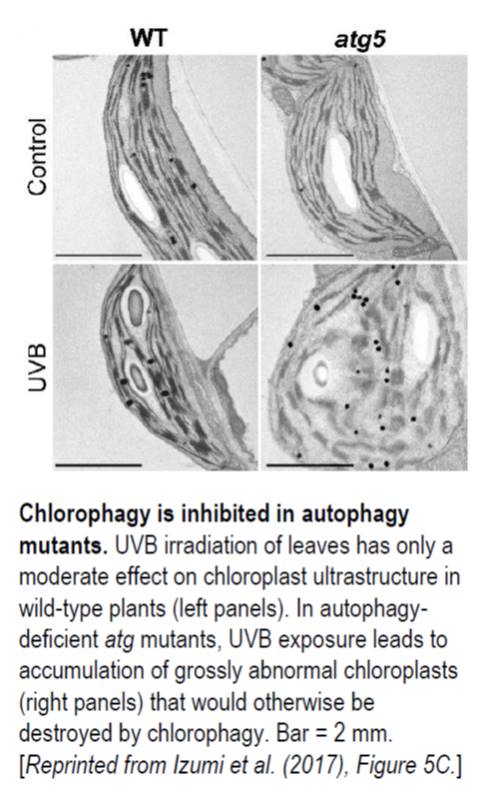
In wild-type leaves, the number of chloroplasts in UVB-exposed leaves decreased approximately 30% by 3 d following exposure. By contrast, no decrease was seen in UVB-exposed atg mutant leaves, which instead accumulated abnormal chloroplasts. In untreated control plants, chloroplast morphology in atg mutant leaves was similar to wild-type morphology (see figure). However, following UVB exposure, the chloroplasts of atg mutant leaves had grossly altered ultrastructure and enhanced ROS production. This strongly suggests that, in normally functioning cells, chlorophagy removes photodamaged chloroplasts following excessive UVB exposure.
The authors showed the involvement of ROS such as H2O2 in the initiation of autophagy following UVB-induced photodamage to chloroplasts. Using concanamycin A, which inhibits the vacuolar H+-ATPase and thus suppresses hydrolytic enzymes, plus an autophagosomal membrane marker, the authors were able to stabilize and visualize tubular autophagosomal structures surrounding individual chloroplasts in the vacuoles of UVB-exposed leaves. Further experiments described in the article showed that strong visible light as well as UVB will induce chlorophagy, and that photodamage-induced chlorophagy involves a pathway distinct from starvation-induced chlorophagy. These findings not only help to elucidate the process of chloroplast turnover in the plant cell but also provide insights into the process of autophagy and organelle turnover in cells.
REFERENCES
Green, D.R., Galluzzi, L., and Kroemer, G. (2011). Mitochondria and the autophagy-inflammation-cell death axis in organismal aging. Science 333: 1109-1112.
Ishida, H., Yoshimoto, K., Izumi, M., Reisen, D., Yano, Y., Makino, A., Ohsumi, Y., Hanson, M.R., and Mae, T. (2008). Mobilization of rubisco and stroma-localized fluorescent proteins of chloroplasts to the vacuole by an ATG gene-dependent autophagic process. Plant Physiol. 148: 142-155.
Izumi, M., Ishida, H., Nakamura, S., and Hidema, J. (2017). Entire photodamaged chloroplasts are transported to the central vacuole by autophagy. Plant Cell 10.1105/tpc.16.00637
Mizushima, N., and Komatsu, M. (2011). Autophagy: renovation of cells and tissues. Cell 147: 728-741.
Nakatogawa, H., Suzuki, K., Kamada, Y., and Ohsumi, Y. (2009). Dynamics and diversity in autophagy mechanisms: lessons from yeast. Nat Rev Mol Cell Bio 10: 458-467
Youle, R.J., and Narendra, D.P. (2011). Mechanisms of mitophagy. Nat Rev Mol Cell Bio 12: 9-14.








Leave a Reply
Want to join the discussion?Feel free to contribute!