No Stakes for High Strength Corn
Kim L Johnson
La Trobe Institute for Agriculture & Food, Department of Animal, Plant and Soil Sciences, La Trobe University, Bundoora, Victoria 3086, Australia
In September 2019 hurricane Dorian swept across to the southeast coast of the USA and strong winds and downpours led to major crop losses in corn and soybean. There is little plants can do in the face of a hurricane, yet it does raise the issue of how crops will stay standing with increasingly variable climates, including storms and high winds, and continual increases in grain yields leading to heavier loads to bear. The need for crops, such as corn, to have strong stalks has never been more apparent. In this issue Jiao et al. (2019) generate corn plants with super strong stems, able to withstand higher forces in mechanical tests. There is a tight correlation between stem strength values and lodging, otherwise known as flattening or breakage of a standing crop, usually by wind or rain in the field (Robertson et al., 2014).
The plant stem can be considered the ‘backbone’ of the plant, supporting the weight of the body and keeping it upright. Anyone with back pain will know that stems must therefore be vulnerable to damage or loss of integrity. The plant’s skeleton is the cell wall, which not only provides incredible strength but is also highly dynamic and plays important roles in plant development. Cells with secondary walls or ‘wood’ are thick, rich in cellulose and often contain lignin. Cells with secondary cell walls are found in vascular tissues and fibres that enable movement of water and nutrients and provide additional strength. Cellulose is the scaffold of plant cell walls, and changes in cellulose in secondary walls influence the mechanical strength of stems, as has been observed in irregular xylem and fragile fibre phenotypes in Arabidopsis, brittle culm in rice, and brittle stalk phenotypes in corn. Investigation of these mutants has revealed key regulators involved in cellulose biosynthesis and secondary wall formation.
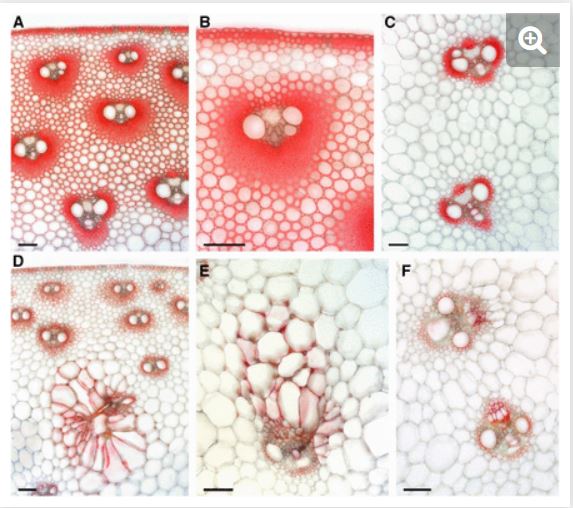
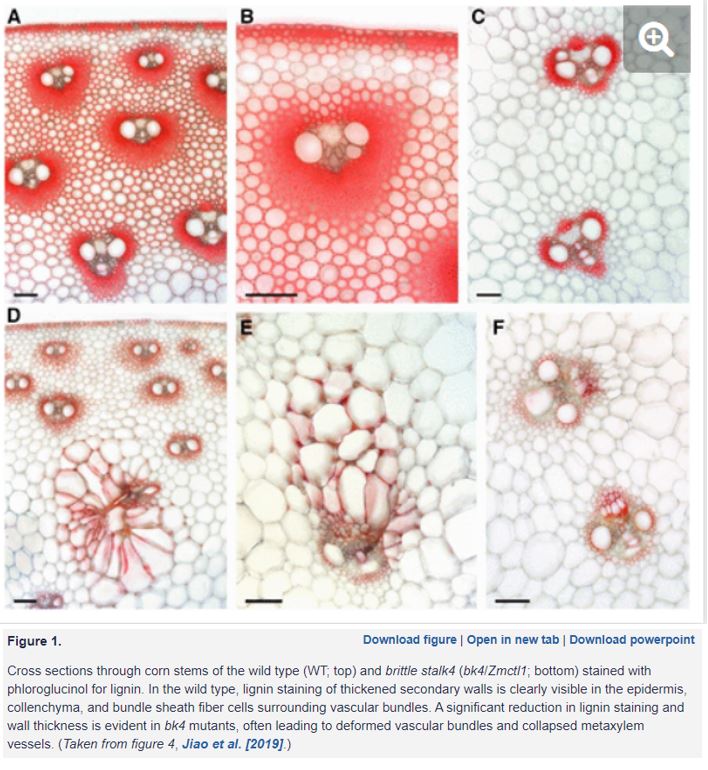 In this issue Jiao et al. (2019) identify brittle stalk4 mutants in corn that show a dwarf phenotype and reduced mechanical strength in stems. Reduced thickness of secondary cell walls is observed in bk4 stems, with dramatic differences in staining of bundle sheath fibre cells surrounding vascular tissue, as well as deformed cell shapes (Figure 1). Along with changes in wall composition these phenotypes point to defects in cell wall biosynthesis/deposition leading to loss of wall integrity. Using a neat PCR-based amplification and cloning approach, a Mutator-insertion was identified in Chitinase-like protein1 (ZmCtl1). Studies of ctl1 mutants in Arabidopsis and rice show similar brittle stem phenotypes, suggesting the role of CTL1 is conserved (Sanchez-Rodriguez et al., 2012; Wu et al., 2012). A member of the glycosyl hydrolase 19 family of enzymes that are proposed to trim carbohydrates, CTL1 itself is unlikely to bind or cleave chitinase as it lacks key catalytic residues (Wu et al., 2012). So how does loss of CTL1 have such a dramatic effect on the cell wall? Decreased cellulose levels are consistently observed in ctl1 mutants along with increases in sugars such as arabinose, xylose and galactose (Sanchez-Rodriguez et al., 2012; Wu et al., 2012; Jiao et al., 2019). Arabidopsis CTL1 can bind glucans, leading to a proposed role for CTL1 as a scaffold protein that influences the cellulose-hemicellulose network (Sanchez-Rodriguez et al., 2012). To explore the relationship with cellulose, bk4/Zmctl1 was crossed to mutants in cellulose synthase genes (ZmCesA10 or ZmCesA11) involved in secondary cell wall formation in corn. Double mutants showed more severe brittle stem phenotypes and reduced height, supporting an association between CTL1 and cellulose (Jiao et al., 2019). When CTL1 was overexpressed in corn, it led to more cellulose and stronger stems; as yet the performance of these plants in the field, with or without wild weather, has not been tested. The biochemical function of CTL1 remains unclear. Determining if CTL1 has hydrolase activity, and if so which carbohydrates it can cleave, would greatly advance our understanding of this fascinating protein. Combined with knowledge of how CTL1 influences the interaction of polysaccharides in secondary walls, we can work towards crops that will keep standing strong.
In this issue Jiao et al. (2019) identify brittle stalk4 mutants in corn that show a dwarf phenotype and reduced mechanical strength in stems. Reduced thickness of secondary cell walls is observed in bk4 stems, with dramatic differences in staining of bundle sheath fibre cells surrounding vascular tissue, as well as deformed cell shapes (Figure 1). Along with changes in wall composition these phenotypes point to defects in cell wall biosynthesis/deposition leading to loss of wall integrity. Using a neat PCR-based amplification and cloning approach, a Mutator-insertion was identified in Chitinase-like protein1 (ZmCtl1). Studies of ctl1 mutants in Arabidopsis and rice show similar brittle stem phenotypes, suggesting the role of CTL1 is conserved (Sanchez-Rodriguez et al., 2012; Wu et al., 2012). A member of the glycosyl hydrolase 19 family of enzymes that are proposed to trim carbohydrates, CTL1 itself is unlikely to bind or cleave chitinase as it lacks key catalytic residues (Wu et al., 2012). So how does loss of CTL1 have such a dramatic effect on the cell wall? Decreased cellulose levels are consistently observed in ctl1 mutants along with increases in sugars such as arabinose, xylose and galactose (Sanchez-Rodriguez et al., 2012; Wu et al., 2012; Jiao et al., 2019). Arabidopsis CTL1 can bind glucans, leading to a proposed role for CTL1 as a scaffold protein that influences the cellulose-hemicellulose network (Sanchez-Rodriguez et al., 2012). To explore the relationship with cellulose, bk4/Zmctl1 was crossed to mutants in cellulose synthase genes (ZmCesA10 or ZmCesA11) involved in secondary cell wall formation in corn. Double mutants showed more severe brittle stem phenotypes and reduced height, supporting an association between CTL1 and cellulose (Jiao et al., 2019). When CTL1 was overexpressed in corn, it led to more cellulose and stronger stems; as yet the performance of these plants in the field, with or without wild weather, has not been tested. The biochemical function of CTL1 remains unclear. Determining if CTL1 has hydrolase activity, and if so which carbohydrates it can cleave, would greatly advance our understanding of this fascinating protein. Combined with knowledge of how CTL1 influences the interaction of polysaccharides in secondary walls, we can work towards crops that will keep standing strong.
References
Jiao S, Hazebroek JP, Chamberlin MA, Perkins M, Sanhu AS, Gupta RK, Simcox KD, Yinghong L, Prall A, Heetland L, Meeley RB, Multani DS (2019) Impairment of a Chitinase-like1 is responsible for the phenotype of a brittle stalk4 mutant in maize. Plant Physiol {https://doi.org/10.1104/pp.19.00615}
Robertson D, Smith S, Gardunia B, Cook D (2014) An Improved Method for Accurate Phenotyping of Corn Stalk Strength. Crop Science 54: 2038-2044
Sanchez-Rodriguez C, Bauer S, Hematy K, Saxe F, Ibanez AB, Vodermaier V, Konlechner C, Sampathkumar A, Ruggeberg M, Aichinger E, Neumetzler L, Burgert I, Somerville C, Hauser MT, Persson S (2012) Chitinase-like1/pom-pom1 and its homolog CTL2 are glucan-interacting proteins important for cellulose biosynthesis in Arabidopsis. Plant Cell 24: 589-607
Wu B, Zhang B, Dai Y, Zhang L, Shang-Guan K, Peng Y, Zhou Y, Zhu Z (2012) Brittle culm15 encodes a membrane-associated chitinase-like protein required for cellulose biosynthesis in rice. Plant Physiol 159: 1440-1452

