Light Triggers the Search for Light
Plants rely on light for photosynthesis. As a consequence, plant growth is strongly shaped by light availability, to optimize photon absorbance in the green tissues. Seedlings that germinated underground grow quickly to escape from the darkness, while plants that are threatened to be overgrown by neighbors rapidly induce stem elongation to win the competition for light. Possibly the most characteristic response of flowering plants to light is the so-called phototropic response, in which plants redirect growth towards the source of light, resulting in bent stems.
Phototropic growth is mostly induced by blue light, which activates two serine/threonine protein kinases: phototropin1 (phot1) and phot2. Activation of the phots at the light-exposed side of the hypocotyl causes dephosphorylation of NON-PHOTOTROPHIC HYPOCOTYL3 (NPH3), which mediates a signaling cascade resulting in a lateral auxin gradient. The high auxin concentrations on the shaded side of the hypocotyl induce cell elongation and the bent phenotype (reviewed in Goyal et al. (2013); Fankhauser and Christie (2015)).
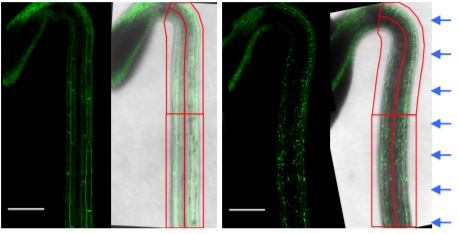
Even though phototropism is a phenotypic adaptation seen in multiple organs and stages of plant development, much of our understanding is based on studies using dark-grown (etiolated) seedlings. In this issue of Plant Physiology, Sullivan et al. (2019) show that phototropism is much stronger in de-etiolated seedlings. They expose dark-grown seedlings to light for eight hours, enough to promote de-etiolation: opening, expansion and greening of cotyledons (Gommers and Monte, 2018). These green seedlings are then returned to darkness for 16 hours and exposed to unilateral blue light to induce phototropism. The authors show that de-etiolation prevents NPH3 dephosphorylation, which results in the enhanced phototropic phenotype. It remains unknown which sites of the NPH3 protein are (de)phosphorylated, how these different phosphorylation states affect the protein’s function, and how this results in auxin redistribution.
The work of Sullivan et al. unravels an interesting interaction between de-etiolation and phototropism. They point out that the difference between light and dark is made at the level of NPH3 phosphorylation. De-etiolation is a complex, irreversible developmental process during which a plant becomes photoautotrophic. Red and blue light wavelengths activate phytochrome (phy) and cryptochrome (cry) photoreceptors, respectively, which deactivate several repressors of de-etiolation: the PHYTOCHROME INTERACTING FACTORS (PIFs) and CONSTITUTIVELY PHOTOMORPHOGENIC1 (COP1). As a consequence, transcription factors that promote de-etiolation, like ELONGATED HYPOCOTYL5 (HY5) and GOLDEN2-LIKE1 (GLK1) and GLK2 are activated. Light-exposed seedlings will stop elongating the hypocotyl, initiate root development and, importantly, expand the cotyledons and develop mature chloroplasts (Gommers and Monte, 2018).
Sullivan et al. find that both phy and cry-mediated de-etiolation delay NPH3 dephosphorylation and promote phototropism, but it remains speculative exactly which players downstream of the photoreceptors are responsible. The pif quadruple (pifq) mutant, which lacks PIF1, PIF3, PIF4 and PIF5, is constitutively photomorphogenic, with a short hypocotyl and open, expanded cotyledons in darkness. This mutant has reduced NPH3 dephosphorylation and a slightly enhanced phototropic phenotype in darkness compared to wild type. Nevertheless, it is still far from as responsive as light-exposed plants, which suggests that, although PIFs repress phototropism in darkness, another light-induced factor promotes the phototrophic response.
The authors discuss that enhanced phototropism in de-etiolated plants helps to maximize their photoautotrophic growth. In other words, it helps to optimize photosynthesis, which makes sense, because phototropism places the leaves towards the sunlight. This, however, makes me wonder if the light-triggered development of etioplasts into functional chloroplasts could help to promote phototropism. It would explain why the dark-grown pifq mutant, constitutively de-etiolated but not green, does not reach the optimum phototrophic response. Chloroplast development is triggered by photoreceptor activation, as chlorophyll synthesis is repressed by PIFs. On the other hand, the plastid releases retrograde signals to inform the nucleus about its developmental state and trigger gene expression to steer chloroplast and whole-plant development (Chan et al., 2016; Hernandez-Verdeja and Strand, 2018). Several recent works have shown that impaired chloroplast development antagonizes light-mediated processes, such as de-etiolation and shade avoidance (Martín et al., 2016; Ortiz-Alcaide et al., 2019). Call it a wild guess, or an interesting hypothesis, but it would be interesting to find out how the greening process itself contributes to phototropism. For now, we can at least conclude that once plants have seen the light, they are more prone to search for it. But to be honest: “Don’t it always seem to go that you don’t know what you got ‘till it’s gone”?
Charlotte M. M. Gommers
Literature Cited
Chan KX, Phua SY, Crisp P, McQuinn R, Pogson BJ (2016) Learning the Languages of the Chloroplast: Retrograde Signaling and Beyond. Annual Review of Plant Biology 67: 25-53
Fankhauser C, Christie John M (2015) Plant Phototropic Growth. Current Biology 25: 384-389
Gommers CMM, Monte E (2018) Seedling establishment: a dimmer switch-regulated process between dark and light signaling. Plant Physiology 176: 1061-1074
Goyal A, Szarzynska B, Fankhauser C (2013) Phototropism: at the crossroads of light-signaling pathways. Trends in Plant Science 18: 393-401
Hernandez-Verdeja T, Strand A (2018) Retrograde signals navigate the path to chloroplast development. Plant Physiology 176: 967-976
Martín G, Leivar P, Ludevid D, Tepperman JM, Quail PH, Monte E (2016) Phytochrome and retrograde signalling pathways converge to antagonistically regulate a light-induced transcriptional network. Nature Communications 7: 11431
Ortiz-Alcaide M, Llamas E, Gomez-Cadenas A, Nagatani A, Martínez-García JF, Rodríguez-Concepción M (2019) Chloroplasts Modulate Elongation Responses to Canopy Shade by Retrograde Pathways Involving HY5 and Abscisic Acid. The Plant Cell 31: 384-398
Sullivan S, Kharshiing E, Laird J, Sakai T, Christie J (2019) De-etiolation Enhances Phototropism by Modulating NON-PHOTOTROPIC HYPOCOTYL 3 Phosphorylation Status. Plant Physiology: pp.00206.02019
Willige BC, Ahlers S, Zourelidou M, Barbosa ICR, Demarsy E, Trevisan M, Davis PA, Roelfsema MRG, Hangarter R, Fankhauser C, Schwechheimer C (2013) D6PK AGCVIII Kinases Are Required for Auxin Transport and Phototropic Hypocotyl Bending in Arabidopsis. The Plant Cell 25: 1674-1688