Cell Wall Remodeling During Wood Development
Madeleine Seale
Department of Plant Sciences, University of Oxford, South Parks Road, Oxford, OX1 3RB, United Kingdom
ORCID: 0000-0002-8924-3943
Wood formation occurs via radial proliferation of vascular cambium cells and the deposition of secondary cell wall layers (Déjardin et al., 2010). This is crucially important for the mechanical stability of the plant and also affects the chemical and mechanical properties of wood, paper, and other ligno-cellulosic products.
An important focus area for cell wall research is to understand better how cell wall composition relates to cell expansion and cell wall mechanical properties. The hemicellulose xyloglucan is one of the major structural components of cell walls and non-covalently bonds with cellulose within cell walls. However, the role of xyloglucans in primary cell walls has been particularly hotly debated as double mutants of the only two xyloglucan synthesis enzymes in Arabidopsis (Arabidopsis thaliana) show fairly mild phenotypes (Cavalier et al., 2008). As a result, some have proposed alternative models for primary cell wall composition suggesting that xyloglucan forms more limited connections with cellulose (Cosgrove 2014). This raises the question as to the function of xyloglucan within cell walls.
In this issue of Plant Physiology, Kushwah and colleagues (2020) investigate the role of xyloglucans in secondary cell wall formation by examining two Arabidopsis xyloglucan remodeling enzymes that are expressed in vascular cambium tissue. Intriguingly, they find that these genes act non-cell autonomously in xylem fiber development.
 When cell walls are remodeled, for example during cell expansion or in response to stress, linkages between cell wall polysaccharides must be undone and reformed. XTHs (XYLOGLUCAN ENDOTRANSGLYCOLSYLASE/HYDROLASEs) are a class of enzymes that perform this function, i.e. they cleave xyloglucan chains and reattach the cleaved end to a new xyloglucan poly- or oligosaccharide (Rose et al., 2002).
When cell walls are remodeled, for example during cell expansion or in response to stress, linkages between cell wall polysaccharides must be undone and reformed. XTHs (XYLOGLUCAN ENDOTRANSGLYCOLSYLASE/HYDROLASEs) are a class of enzymes that perform this function, i.e. they cleave xyloglucan chains and reattach the cleaved end to a new xyloglucan poly- or oligosaccharide (Rose et al., 2002).
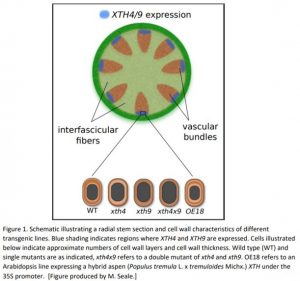
In aspen (Populus tremula), several XTHs have been previously identified that are expressed in vascular cambium (Sundell et al., 2017). To study their function in greater detail, the authors selected 2 Arabidopsis XTHs (XTH4 and 9) based on sequence similarity to the aspen genes and based on expression pattern (Fig 1).
Using histology, spectroscopy and enzymatic assays, the authors investigated a range of phenotypes in Arabidopsis lines with mutations in the two XTH genes and in a line overexpressing an aspen XTH. They found that the two xth mutants exhibited reduced xyloglucan endotransglycosylase activity, while the overexpressor line showed an increase. Furthermore, the mutants had increased hypocotyl diameters, but the overexpressor had narrower hypocotyls.
Not all phenotypes showed the straightforward opposite effects in mutants versus overexpressor. For example, plant height was greater than wild type in all mutants as well as the overexpressor and the number of cell wall layers showed interesting tissue-dependent effects that varied with the single xth mutants, the double mutant and the overexpressor (Fig 1). The authors neatly demonstrated that the number of cell wall layers increased with increasing cellulose content but not with lignin content. Furthermore, changes to lignin content in the xth mutants compared to wild type were clearly observed in interfascicular fibres. Given that XTHs are not expressed in these tissues, it appears that XTHs act non-cell autonomously.
A particularly interesting finding was that XTH4 and XTH9 appear to regulate vessel fiber cell length, primarily by affecting intrusive growth. XTHs have not previously been associated with intrusive growth, so this represents an important area for future research.
The impact of altered xyloglucan endotransglycosylase activity on intrusive growth and the non-cell autonomous phenotypes indicated that structural or mechanical features may be altered compared to wild type plants. For other Arabidopsis XTHs, overexpression has led to altered mechanical properties and thicker cell walls that were thought to be an indirect result of increased cellulose deposition (Miedes et al., 2013). To gain further insight into possible indirect effects of modified XTH expression on cell wall properties, Kushwah et al. (2020) investigated the expression of genes involved in cell wall integrity sensing. They found that genes encoding THESEUS, FERONIA and WALL-ASSOCIATED KINASES all exhibited altered expression in either the xth mutants or the XTH overexpressor. This indicates that cell wall integrity signaling may mediate some of the phenotypes observed.
The authors have identified some key functions of XTH4 and XTH9 and established their role in regulating xyloglucan within vascular cambium. Additionally, they have investigated more wide-reaching implications of altering xyloglucan linkages and highlighted the complex relationships between XTHs and cell walls, potentially reflecting feedback between cell wall deposition, composition and remodeling in within and between tissue types. It will be interesting in the future to understand further how intrusive fiber growth is regulated by XTH activity and the mechanism behind the non-cell autonomous effects on cell walls. Other recent work in the shoot apical meristem found that functional microtubule dynamics were particularly important when xyloglucans were perturbed (Zhao et al., 2019). This suggests that examination of the cytoskeleton in vascular cambium may be fruitful for understanding the interactions between XTHs, xyloglucans and other cell wall components.
LITERATURE CITED
Cavalier DM, Lerouxel O, Neumetzler L, Yamauchi K, Reinecke A, Freshour G, Zabotina OA, Hahn MG, Burgert, I, Pauly M et al. (2008) Disrupting Two Arabidopsis thaliana Xylosyltransferase Genes Results in Plants Deficient in Xyloglucan, a Major Primary Cell Wall Component. Plant Cell, 20: 1519–1537
Cosgrove D (2014) Re-constructing our models of cellulose and primary cell wall assembly. Current Opinion in Plant Biology 22: 122-131
Déjardin A, Laurans F, Arnaud D, Breton C, Pilate G, Leplé J-C (2010) Wood formation in Angiosperms. Comptes Rendus Biologies, 333: 325-334
Kushwah S, Banasiak A, Nishikubo N, Derba-Maceluch M, Majda M, Endo S, Kumar V, Gomez L, Gorzsas A, McQueen-Mason S et al. (2020) Arabidopsis XTH4 and XTH9 contribute to wood cell expansion and secondary wall formation. Plant Physiology https://doi.org/10.1104/pp.19.01529
Rose JKC, Braam J, Fry SC, Nishitani K (2002) The XTH Family of Enzymes Involved in Xyloglucan Endotransglucosylation and Endohydrolysis: Current Perspectives and a New Unifying Nomenclature. Plant and Cell Physiology 43: 1421–1435
Miedes E, Suslov D, Vandenbussche F, Kenobi K, Ivakov A, van der Straeten D, Lorences EP, Mellerowicz EJ, Verbelen J-P, Vissenberg K (2013) Xyloglucan endotransglucosylase/hydrolase (XTH) overexpression affects growth and cell wall mechanics in etiolated Arabidopsis hypocotyls. Journal of Experimental Botany 64:2481–2497
Sundell D, Street NR, Kumar M, Mellerowicz EJ, Kucukoglu M, Johnsson C, Kumar V, Mannapperuma C, Delhomme N, Nilsson O et al. (2017) AspWood: High-Spatial-Resolution Transcriptome Profiles Reveal Uncharacterized Modularity of Wood Formation in Populus tremula. Plant Cell 29: 1585-1604
Zhao F, Chen W, Sechet J, Martin M, Bovio S, Lionnet C, Long Y, Battu V, Mouille G, Monéger F et al. (2019) Xyloglucans and Microtubules Synergistically Maintain Meristem Geometry and Phyllotaxis. Plant Physiology 181: 1191-1206







