A Novel Role of Ring Chromosomes as Evolutionary Drivers of Herbicide Resistance
In eukaryotes, chromosomes are linear structures; in contrast, a ring-shaped chromosome is a rare structure that results from the fusion of broken ends of linear chromosomes. Ring chromosomes may be induced by radiation or may occur spontaneously. In humans, ring chromosomes occur in approximately 1:50,000 embryos, and often are associated with growth deficiency and disease. Cancer cells are likely to produce ring chromosomes due to abnormal cell cycles (Pristyazhnyuk and Menzorov, 2017). Ring chromosomes have also been observed in several plant species (Singh, 2016). However, the functional significance and selective value of ring chromosomes in plants were not known. In this issue, Koo et al. (2018) report a novel role of a ring chromosome as an evolutionary driver of herbicide resistance in common waterhemp (Amaranthus tuberculatus).
Ring chromosomes were first observed in Drosophila melanogaster (Morgan, 1926) and maize (Zea mays; McClintock, 1931). Using x-rays, McClintock (1931, 1932, 1938) produced and identified ring chromosomes that resulted in variegation in the stalk and leaves of mutagenized maize. One variegation type, which shows brown stripes along the veins of the leaves and leaf sheaths, is associated with the occurrence of a ring chromosome that carries the locus Brown midrib (Bm1). The presence of the Bm1 allele on the ring chromosome results in a normal green color; loss of this ring chromosome results in brown variegation due to the presence of the recessive bm1 allele on the rod chromosome (McClintock, 1932, 1938). The ring chromosome is probably derived from a broken fragment of a normal chromosome induced by x-ray treatment. Due to the instability of broken chromosomes lacking the protection afforded by telomeres, broken ends fuse to form a ring-shaped chromosome (Fig. 1A). If both the deficient rod chromosome and the ring chromosome carry part of a functional centromere, they will be maintained in the daughter cells after cell division (McClintock, 1932, 1938).
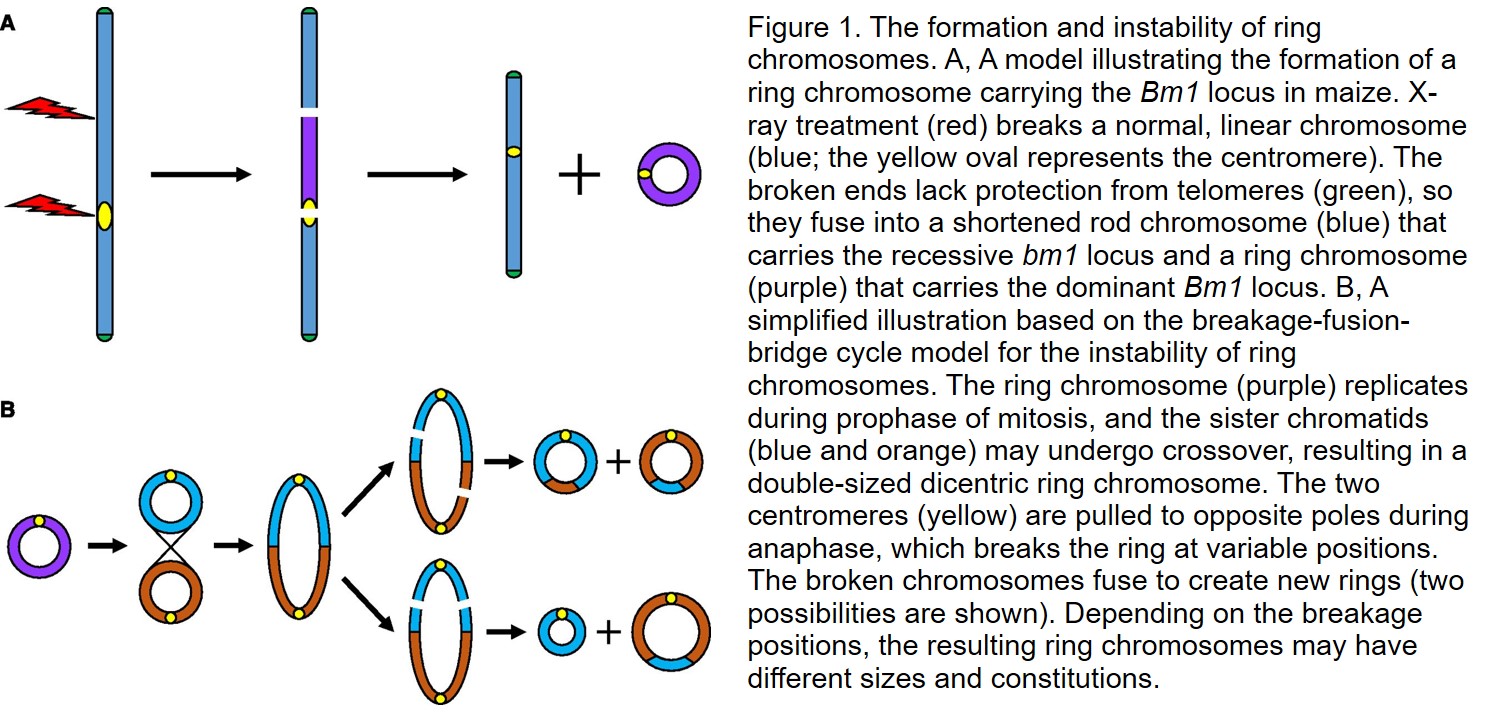
McClintock (1938) further showed that ring chromosomes are not stable and can change size and constitution, or can be lost during mitotic cycles, leading her to propose the breakage-fusion-bridge cycle model (McClintock, 1941a, 1941b). In this model, sister chromatid exchange may result in a double-sized ring chromosome with two centromeres, which are pulled to opposite spindle poles during anaphase. The fused ring chromosome (bridge) ruptures, and the broken ends fuse to form new ring chromosomes. Because the rupture occurs at variable positions, the resulting rings vary in size (Fig. 1B). If a ring chromosome stays at the equatorial plate during anaphase, it may not enter either daughter cell and may subsequently be eliminated after the cell cycle (McClintock, 1941a, 1941b).
Despite the instability of these ring chromosomes, a new study in this issue shows that they can be retained under selection. In this study, Koo et al. (2018) demonstrate retention of a ring chromosome under intense herbicide selection in waterhemp. Waterhemp is a notorious weed that grows in maize and soybean (Glycine max) fields, and has become resistant to several herbicides, including glyphosate (N-(phosphonomethyl)glycine), known commercially as Roundup. Glyphosate interacts with and inhibits the enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), which is required for the biosynthesis of aromatic amino acids (Koo et al., 2018). In an earlier report, Dillon et al. (2017) showed that increased copies of the EPSPS gene in the waterhemp genome correlated with increased transcript level of EPSPS and a marked resistance to glyphosate.
Interestingly, the authors observed an additional chromosome harboring several copies of EPSPS in two highly glyphosate-resistant individuals, resulting in 2n = 32 + 1 chromosomes (Dillon et al., 2017). Now Koo et al. (2018) use 4′,6-diamidino-2-phenylindole staining and fluorescence in situ hybridization to reveal that this extra chromosome is a ring chromosome. The ring chromosome can be sexually transmitted to the next generation, indicating that it has a centromere. Consistent with McClintock’s observations (McClintock, 1938), the waterhemp ring chromosomes varied in size and copy number in the progenies of crosses between resistant and susceptible plants. In a surprising twist, while the authors expected the typical EPSPS signals in both the ring and rod chromosomes inherited from the parents, they observed additional EPSPS signals in rod chromosomes in the cells that harbor the ring chromosomes. The authors propose that these extra EPSPS copies on the rod chromosomes may have translocated from ring chromosomes through ectopic recombination with nonhomologous chromosomes (Koo et al., 2018).
The occurrence of a ring chromosome harboring EPSPS in waterhemp is likely to be associated with the high selection pressure imposed by spraying with glyphosate. Moreover, the fact that functional genes may translocate from ring chromosomes onto rod chromosomes provides a potential mechanism for rapid evolution under extreme environments. The deciphering of this mechanism opens a new avenue for studying chromosome engineering and genomic modification in plants.
REFERENCES
Dillon A, Varanasi VK, Danilova TV, Koo D-H, Nakka S, Peterson DE, Tranel PJ, Friebe B, Gill BS, Jugulam M (2017) Physical mapping of amplified copies of the 5-enolpyruvylshikimate-3-phosphate synthase gene in glyphosate-resistant Amaranthus tuberculatus. Plant Physiol 173: 1226–1234
Koo D-H, Jugulam M, Putta K, Cuvaca IB, Peterson DE, Currie RS, Friebe B, Gill BS (2018) Gene duplication and aneuploidy trigger rapid evolution of herbicide resistance in common waterhemp. Plant Physiol 176: 1932–1938
McClintock B (1931) Cytological observations of deficiencies involving known genes, translocations and an inversion in Zea mays. Missouri Agricultural Experiment Station Research Bulletin 163: 3–30
McClintock B (1932) A correlation of ring-shaped chromosomes with variegation in Zea mays. Proc Natl Acad Sci USA 18: 677–681
McClintock B (1938) The production of homozygous deficient tissues with mutant characteristics by means of the aberrant mitotic behavior of ring-shaped chromosomes. Genetics 23: 315–376
McClintock B (1941a) The association of mutants with homozygous deficiencies in Zea mays. Genetics 26: 542–571
McClintock B (1941b) The stability of broken ends of chromosomes in Zea mays. Genetics 26: 234–282
Morgan LV (1926) Correlation between shape and behavior of a chromosome. Proc Natl Acad Sci USA 12: 180–181
Pristyazhnyuk IE, Menzorov AG (2017) Ring chromosomes: from formation to clinical potential. Protoplasma https://doi.org/10.1007/s00709…
Singh RJ (2016) Chromosome abbreviations. In RJ Singh, ed, Plant Cytogenetics, Ed 3. CRC Press, Boca Raton, FL, pp 161–163
Published March 2018. DOI: https://doi.org/10.1104/pp.18.00083