A GDSL lipase is required for anther and pollen development
Department of Plant Pathology, Kansas State University, 1712 Claflin Road, Throckmorton Hall, Manhattan, KS, 66506, USA
Male fertility in angiosperms requires specific spatiotemporal gene expression patterns and cellular differentiation during male reproductive development (Wilson and Zhang, 2009; Lin et al., 2017). The formation of the male gametophyte begins inside the anther with the formation of a large microspore mother cell, which eventually forms into individual microgametophytes (pollen grains). Initially, microgametophytes rely on nutrients and molecules arising from the coordinated degradation of the tapetum, the innermost layer of the anther (Li et al., 2006). Later, pollen maturation requires nutrients provided by starch and/or lipids. However, the genes encoding lipid-modifying enzymes such as lipases and esterases and the associated regulatory mechanisms in plant male fertility remain elusive.
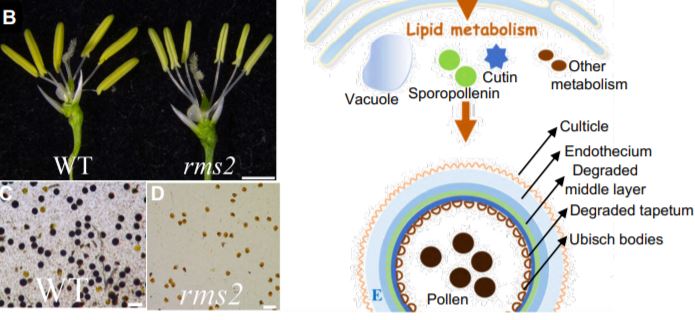
 In this issue of Plant Physiology, Zhao et al. (2020) characterized a gene, Rice Male Sterile 2 (RMS2, encoding a GDSL lipase protein), that is required for anther development and pollen fertility by affecting tapetal degradation, the formation of anther cuticle and pollen exine, and central vacuole development (Zhao et al., 2020). Deletions and substitution at RMS2 resulted in rice male sterility, with abnormal anther and pollen phenotypes. The authors first screened a rice mutant population and identified a mutant, rice male sterile 2 (rms2), with a male sterility phenotype, including delayed heading dates; partly sheathed panicles; conspicuous, shrunken, and white anthers; and abnormal pollen grains, but with no effect on female fertility (Figure 1A to 1D) (Zhao et al., 2020).
In this issue of Plant Physiology, Zhao et al. (2020) characterized a gene, Rice Male Sterile 2 (RMS2, encoding a GDSL lipase protein), that is required for anther development and pollen fertility by affecting tapetal degradation, the formation of anther cuticle and pollen exine, and central vacuole development (Zhao et al., 2020). Deletions and substitution at RMS2 resulted in rice male sterility, with abnormal anther and pollen phenotypes. The authors first screened a rice mutant population and identified a mutant, rice male sterile 2 (rms2), with a male sterility phenotype, including delayed heading dates; partly sheathed panicles; conspicuous, shrunken, and white anthers; and abnormal pollen grains, but with no effect on female fertility (Figure 1A to 1D) (Zhao et al., 2020).
To identify the molecular mechanism underlying pollen failure in the mutant, the authors carried out a histological analysis of developing rms2 pollen. rms2 mutant pollen can successfully go through the early stages of development, but becomes abnormal at the start of the vacuolated pollen stage prior to microgametogenesis. Compared with the wild type, the authors didn’t observe the large vacuolated microspores but found an intact tapetum and no sign of degradation at this stage just prior to microspore mitosis I. Furthermore, the delayed synthesis of anther cuticle and pollen exine observed in the mutant, coincides with the delayed degradation of the tapetum. Thus, pollen maturation probably fails in the rms2 mutant due to a disruption of nutrient filling that is supported by proper degradation of the tapetum.
The authors found that the rms2 phenotype is due to a single recessive nuclear gene, which they mapped and identified as encoding a GDSL lipase. Spatiotemporal expression analysis showed that RMS2 is predominantly transcribed in the anther during the early stages of pollen development. By screening the interactions with key male-fertility regulators, the groups revealed that RMS2 is targeted by at least by two transcription factors, including Undeveloped Tapetum 1 (UDT1) and Persistent Tapetal Cell 1 (PTC1), which serve as essential regulators during early tapetum development.
By conducting a GFP-labeled assay, the group showed that RMS2 is localized to the endoplasmic reticulum (ER). Based on the annotated GDSL domain, the authors hypothesized that RMS2 may possess hydrolase activity and function in lipid metabolism. However, rice RMS2 clusters far away from other GDSL proteins with diverse roles in plant growth and defense and a recently identified maize protein ZmMs30 that causes maize male sterility, suggesting RMS2 has distinct roles in rice male fertility (Lee et al., 2009; Kikuta et al., 2012; Gao et al., 2017; An et al., 2019; Watkins et al., 2019).
Although recombinant RMS2 did not exhibit hydrolase activity in an in vitro assay, total anther proteins isolated from rms2 had reduced hydrolytic activity, suggesting that RMS2 has general lipase activity. Furthermore, the group conducted a comparative metabolomic analysis and established that the metabolic profiles of rms2 pollen and anthers was strikingly different from those of wild-type samples. In addition to revealing differences in lipid metabolites, metabolomic analysis also revealed altered levels of several plant hormones, including gibberellin, auxin, and jasmonic acid. The extensive changes in primary and secondary metabolic pathways suggest that RMS2 may be involved not only in lipid metabolism but also in other unknown pathways that regulate male fertility.
The authors present a model showing how RMS2 contributes to anther and pollen development in rice (Figure 1E). When male-fertility transcriptional regulators bind to the RMS2 promoter, the gene is expressed predominantly in the developing anther. The encoded ER-localized GDSL lipase functions during pollen mitosis to control tapetal degradation, the formation of anther cuticle, pollen exine, and the central vacuole. All of these events are required for pollen development and maturation. The functions of RMS2 in rice male sterility make it a promising genetic resource for hybrid rice breeding. Further research is needed to investigate whether the mutation in RMS2 affects other important agronomic traits.
References
An, X., Dong, Z., Tian, Y., Xie, K., Wu, S., Zhu, T., Zhang, D., Zhou, Y., Niu, C., Ma, B., Hou, Q., Bao, J., Zhang, S., Li, Z., Wang, Y., Yan, T., Sun, X., Zhang, Y., Li, J., and Wan, X. (2019). ZmMs30 Encoding a Novel GDSL Lipase Is Essential for Male Fertility and Valuable for Hybrid Breeding in Maize. Mol Plant 12, 343-359.
Gao, M., Yin, X., Yang, W., Lam, S.M., Tong, X., Liu, J., Wang, X., Li, Q., Shui, G., and He, Z. (2017). GDSL lipases modulate immunity through lipid homeostasis in rice. PLoS Pathog 13, e1006724.
Kikuta, Y., Ueda, H., Takahashi, M., Mitsumori, T., Yamada, G., Sakamori, K., Takeda, K., Furutani, S., Nakayama, K., Katsuda, Y., Hatanaka, A., and Matsuda, K. (2012). Identification and characterization of a GDSL lipase-like protein that catalyzes the ester-forming reaction for pyrethrin biosynthesis in Tanacetum cinerariifolium- a new target for plant protection. Plant J 71, 183-193.
Lee, D.S., Kim, B.K., Kwon, S.J., Jin, H.C., and Park, O.K. (2009). Arabidopsis GDSL lipase 2 plays a role in pathogen defense via negative regulation of auxin signaling. Biochem Biophys Res Commun 379, 1038-1042.
Li, N., Zhang, D.S., Liu, H.S., Yin, C.S., Li, X.X., Liang, W.Q., Yuan, Z., Xu, B., Chu, H.W., Wang, J., Wen, T.Q., Huang, H., Luo, D., Ma, H., and Zhang, D.B. (2006). The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development. Plant Cell 18, 2999-3014.
Lin, H., Yu, J., Pearce, S.P., Zhang, D., and Wilson, Z.A. (2017). RiceAntherNet: a gene co-expression network for identifying anther and pollen development genes. Plant J 92, 1076-1091.
Watkins, J.L., Li, M., McQuinn, R.P., Chan, K.X., McFarlane, H.E., Ermakova, M., Furbank, R.T., Mares, D., Dong, C., Chalmers, K.J., Sharp, P., Mather, D.E., and Pogson, B.J. (2019). A GDSL Esterase/Lipase Catalyzes the Esterification of Lutein in Bread Wheat. Plant Cell 31, 3092-3112.
Wilson, Z.A., and Zhang, D.B. (2009). From Arabidopsis to rice: pathways in pollen development. J Exp Bot 60, 1479-1492.