Turning up the heat: the alternative oxidase pathway drives thermogenesis in cycad cones
Kim L Johnson
La Trobe Institute for Agriculture & Food, Department of Animal, Plant and Soil Sciences, La Trobe University, Bundoora, Victoria 3086, Australia
Flowers are mostly associated with pleasant smells; the sweet fragrance of a rose, the exotic scent of jasmine, the fresh smell of spring blossom. Some flowers, however, produce quite the opposite, releasing foul smelling odours that attract a more discerning class of pollinators. One of the most famous examples is Raffelesia, also known as the corpse flower; it produces the world’s largest flower and has the heady scent of rotting meat. To release these volatile odours, the flower heats up in a process called thermogenesis. This incredible process can result in heating the flower up to 10°C above the surrounding air temperature. The amazing ability of the plant to heat to such temperatures for the mutual benefit of pollinator development and plant reproduction shows the ingenuity of plant co-evolution. Raffelesia is just one of a number of plants with this ability. Thermogenesis of the reproductive organs can occur in inflorescences, flowers or cones of species from both gymnosperms and angiosperms. Approximately half of all known thermogenic species are cycads, where the male cones can heat for weeks on end (Roemer et al., 2013; Ito-Inaba et al., 2019). This is unlike many angiosperm thermogenic species, which heat for only hours to days (Miller et al., 2011).
So how do plants turn up the heat? The answer lies in the mitochondria, the energy factory of the cell. In addition to the respiratory pathway that produces energy in the form of ATP, an alternative pathway exists that is catalysed by alternative oxidase (AOX) and does not produce ATP. First identified as a distinct oxidase pathway resistant to inhibition by the poison cyanide, roles for AOX include preventing formation of reactive oxygen species, and in angiosperms, regulation of floral thermogenesis.
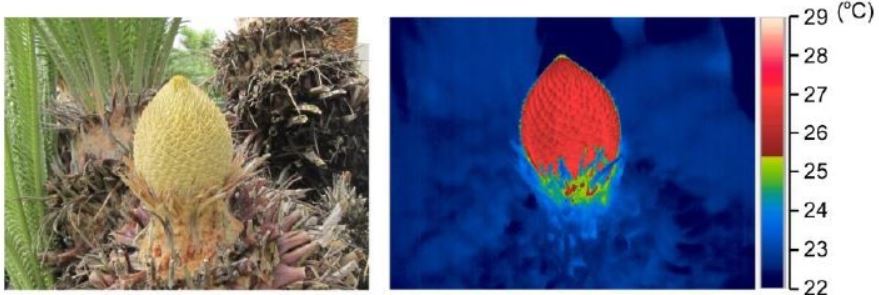
In gymnosperms the role of AOX in thermogenesis has remained unclear. In this issue of Plant Physiology, Ito-Inaba et al. (2019) investigate cone thermogenesis in Cycad revoluta and not only identify a role for AOX, but discover intriguing mitochondrial structures in thermogenic tissue. Male cones in Cycad revoluta can heat to an incredible 11.5°C above air temperature (see figure). The cones consist of a stunning spiral array of microsporophylls with the microsporangia, in which the pollen grains develop, attached to the surface. The microsporophylls are proposed to be thermogenic and Ito-Inaba et al. (2019) identify distinct mitochondrial structures in this tissue, including huge mitochondria in the epidermal cells that are suggested to relate to heat production. Mitochondria extracted from microsporophylls were also shown to have greater AOX protein levels and increased capacity for cyanide-resistant respiration than those in the non-thermogenic microsporangia. Although a number of AOX gene sequences have now been identified in gymnosperms (Neimanis et al., 2013), which genes are associated with thermogenesis is unknown. In a neat molecular study, two AOX genes were identified in Cycad revoluta. Higher levels of CrAOX1 transcripts in microsporophylls suggest this encodes the AOX responsible for male cone thermogenesis.
The use of the AOX pathway for floral thermogenesis raises questions of the specificity and regulation of such processes. AOX proteins can be differentially regulated at the transcriptional and post-translational levels, yet the molecular mechanisms of the different isoforms remain to be fully understood (Selinski et al., 2018). The significance of the distinct mitochondrial structures associated with thermogenic tissue in Cycad revoluta is unclear and opens many avenues to be explored. Are they required to accommodate changes in respiration and increased AOX capacity? Thermogenesis is a fascinating plant innovation for odour-based communication and an area where we can certainly sniff-out more information.
 Figure
Figure
Figure 1. Thermogenesis in a Cycas revoluta male cone showing heating up to 6°C above night time temperatures (22°C). Adapted from Ito-Inaba et al., 2019.
LITERATURE CITED
Ito-Inaba Y, Sato M, Sato MP, Kurayama Y, Yamamoto H, Ohata M, Ogura Y, Hayashi T, Toyooka K, Inaba T (2019) Alternative oxidase capacity of mitochondria in microsporophylls may function in cycad thermogenesis. Plant Physiol
Miller RE, Grant NM, Giles L, Ribas-Carbo M, Berry JA, Watling JR, Robinson SA (2011) In the heat of the night–alternative pathway respiration drives thermogenesis in Philodendron bipinnatifidum. New Phytol 189: 1013-1026
Neimanis K, Staples JF, Huner NP, McDonald AE (2013) Identification, expression, and taxonomic distribution of alternative oxidases in non-angiosperm plants. Gene 526: 275-286
Roemer RB, Terry LI, Marler TE (2013) Cone thermogenesis and its limits in the tropical Cycas micronesica (Cycadaceae): association with cone growth, dehiscence, and post-dehiscence phases. Am J Bot 100: 1981-1990
Selinski J, Hartmann A, Deckers-Hebestreit G, Day DA, Whelan J, Scheibe R (2018) Alternative Oxidase Isoforms Are Differentially Activated by Tricarboxylic Acid Cycle Intermediates. Plant Physiol 176: 1423-1432