Plastid Sulfate Transporters Open Doors To Abiotic Stress Resistance
Climate change is causing more extreme weather patterns and increasing challenges for agriculture. The unusually dry European summer of 2018 caused an 8% drop in cereal production (source: https://ec.europa.eu/info/sites/info/files/food-farming-fisheries/farming/documents/short-term-outlook-autumn-2018_en.pdf). Plants need water for photosynthesis and to maintain turgor pressure; drought-stressed plants wilt, turn yellow, and eventually die. Stomata quickly close during drought stress, limiting water loss by transpiration. Since drought is first perceived by roots, this response requires a signal to move rapidly upward to the leaves, where stomata are localized.
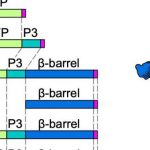
The anion sulfate (SO4–2) quickly accumulates in the xylem upon drought stress and is therefore thought to fulfill this role as a signaling molecule (Malcheska et al., 2017). Several classes of SO4–2 transporters (SULTRs) are responsible for SO4–2 uptake from the soil (root-localized SULTR1;1 and SULTR1;2) and transport into the xylem and towards the shoot (SULTR2;1 and SULTR2;2) or into the phloem and back to the roots (SULTR1;3) (figure). At the cellular level, SULTR4;1 and SULT4;2 mediate SO4–2 efflux from the vacuole, where it is stored. The SULTR3 class of transporters has been less well studied, although SULTR3;1 was shown to localize to the chloroplast membrane and import SO4–2 into these organelles (Cao et al., 2014). In this issue of Plant Physiology, Chen and colleagues (2019) characterize the function of all five SULTR3 transporters in Arabidopsis thaliana. They confirm that all are localized to the chloroplast membrane and facilitate the import of SO4–2 into the plastids (figure).
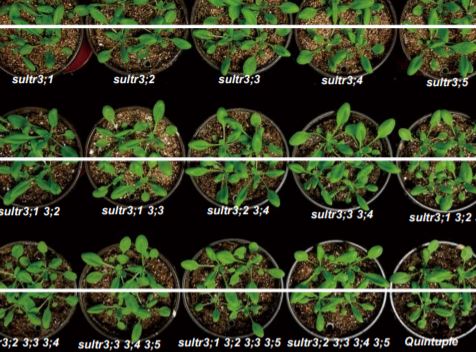
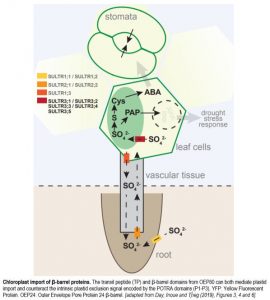 SO4–2 accumulation in the chloroplasts is essential, as is supported by the impaired-growth phenotype of the sultr3-quintuple mutant that Chen et al. created for their study. SO4–2 is the main source of sulfur, which is incorporated into the amino acids cysteine and methionine, via a plastid-localized metabolic process. Cysteine is essential for ABA biosynthesis (figure). Despite the limited nutrient uptake from the soil during drought, SO4–2 quickly accumulates in the xylem sap, is transported towards green tissues, is imported into the chloroplasts, and ultimately causes an increase in ABA and stomatal closure (Malcheska et al., 2017; Batool et al., 2018). Chen et al. now show that this rapid drought response depends on all five SULTR3 transporters, resulting in an 80% reduction in ABA levels in the quintuple mutant compared to wild type plants.
SO4–2 accumulation in the chloroplasts is essential, as is supported by the impaired-growth phenotype of the sultr3-quintuple mutant that Chen et al. created for their study. SO4–2 is the main source of sulfur, which is incorporated into the amino acids cysteine and methionine, via a plastid-localized metabolic process. Cysteine is essential for ABA biosynthesis (figure). Despite the limited nutrient uptake from the soil during drought, SO4–2 quickly accumulates in the xylem sap, is transported towards green tissues, is imported into the chloroplasts, and ultimately causes an increase in ABA and stomatal closure (Malcheska et al., 2017; Batool et al., 2018). Chen et al. now show that this rapid drought response depends on all five SULTR3 transporters, resulting in an 80% reduction in ABA levels in the quintuple mutant compared to wild type plants.
SO4–2 uptake into the chloroplasts also contributes to drought stress tolerance in an ABA-independent pathway. During secondary sulfur metabolism in the plastids, an important byproduct accumulates: 3’-phosphoadenosine 5’-phosphate (PAP). PAP can be degraded to AMP by the Arabidopsis 3’(2’),5’-bisphosphate nucleotidase (SAL1) enzyme, but can also move out of the chloroplast. PAP is a retrograde signal, able to enter the nucleus, where it inhibits the RNA-degrading ability of several 5’ to 3’ exoribonucleases (XRNs). This results in the accumulation of drought-related gene products and increased stress tolerance (figure). As a proof of concept, sal1 mutants of Arabidopsis accumulate PAP and are able to withstand drought much better than wild-type plants (Wilson et al., 2009; Estavillo et al., 2011; Chan et al., 2013).
The data provided by Chen et al. confirm that chloroplasts play a key role during drought stress adaptation. Although far away from the source of the stress, chloroplasts gather information and release the signals to undertake action. Chloroplasts are the photosynthetic and metabolic factories but, as supported by an increasing number of studies, also act as abiotic stress signaling modules.
Literature Cited
Batool S, Uslu VV, Rajab H, Ahmad N, Waadt R, Geiger D, Malagoli M, Xiang C-B, Hedrich R, Rennenberg H, Herschbach C, Hell R, Wirtz M (2018) Sulfate is Incorporated into Cysteine to Trigger ABA Production and Stomatal Closure. The Plant Cell 30: 2973-2987
Cao M-J, Wang Z, Zhao Q, Mao J-L, Speiser A, Wirtz M, Hell R, Zhu J-K, Xiang C-B (2014) Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana. The Plant Journal 77: 604-615
Chan KX, Wirtz M, Phua SY, Estavillo GM, Pogson BJ (2013) Balancing metabolites in drought: the sulfur assimilation conundrum. Trends in Plant Science 18: 18-29
Chen Z, Zhao P-X, Miao Z-Q, Qi G-F, Wang Z, Yuan Y, Ahmad N, Cao M-J, Hell R, Wirtz M, Xiang C-B (2019) SULTR3s involve in chloroplast sulfate uptake and affects ABA biosynthesis and stress response. Plant Physiology
Estavillo GM, Crisp PA, Pornsiriwong W, Wirtz M, Collinge D, Carrie C, Giraud E, Whelan J, David P, Marin E, Pogson BJ (2011) Evidence for a SAL1-PAP Chloroplast Retrograde Pathway That Functions in Drought and High Light Signaling in Arabidopsis. The Plant Cell 23: 3992-4012
Malcheska F, Ahmad A, Batool S, Müller HM, Ludwig-Müller J, Kreuzwieser J, Randewig D, Hänsch R, Mendel RR, Hell R, Wirtz M, Geiger D, Ache P, Hedrich R, Herschbach C, Rennenberg H (2017) Drought-Enhanced Xylem Sap Sulfate Closes Stomata by Affecting ALMT12 and Guard Cell ABA Synthesis. Plant Physiology 174: 798-814
Wilson PB, Estavillo GM, Field KJ, Pornsiriwong W, Carroll AJ, Howell KA, Woo NS, Lake JA, Smith SM, Harvey Millar A, Von Caemmerer S, Pogson BJ (2009) The nucleotidase/phosphatase SAL1 is a negative regulator of drought tolerance in Arabidopsis. Plant Journal 58: 299-317