Plant Science Research Weekly, September 12, 2025
Review: Plant receptor-like kinases sense extrinsic and host-derived signals and shape the microbiome
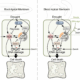
 Plant genomes contain hundreds of genes encoding receptor-like kinases (RLKs), which are membrane-spanning proteins with an extracellular perception domain and an intracellular signaling domain. Many RLKs have roles in identifying surface proteins of pathogens, some recognize specific signals from mutualists such as rhizobia, and others recognize small molecules of various endogenous and exogenous origins. In this new review by Bhat and Haney, the authors explore how RLKs shape the microbiome through perception and signaling. Several RLKs have very narrow species-specific recognition functions, and its unlikely that these have much role in shaping the microbiome. Rather, RLKs that perceive plant cellular responses or physiological signals might have more important roles. As an example, in response to low-nutrient stress plants secrete C-terminally-encoded peptides (CEPs) that are perceived by RLKs and initiate recruitment of beneficial microbes. Similarly, stressed plants might produce signals perceived by differnt RLKs leading to a restructuring of the microbiota; this model is supported by mutants of the FERONIA (FER) RLK that show altered microbiota composition. Activated RLKs can lead to altered calcium fluxes and ROS production as well as transcriptomic changes, any and all of which can contribute to altered microbiota. (Summary by Mary Williams @PlantTeaching.bsky.social) Cell Host Microbe 10.1016/j.chom.2025.07.012
Plant genomes contain hundreds of genes encoding receptor-like kinases (RLKs), which are membrane-spanning proteins with an extracellular perception domain and an intracellular signaling domain. Many RLKs have roles in identifying surface proteins of pathogens, some recognize specific signals from mutualists such as rhizobia, and others recognize small molecules of various endogenous and exogenous origins. In this new review by Bhat and Haney, the authors explore how RLKs shape the microbiome through perception and signaling. Several RLKs have very narrow species-specific recognition functions, and its unlikely that these have much role in shaping the microbiome. Rather, RLKs that perceive plant cellular responses or physiological signals might have more important roles. As an example, in response to low-nutrient stress plants secrete C-terminally-encoded peptides (CEPs) that are perceived by RLKs and initiate recruitment of beneficial microbes. Similarly, stressed plants might produce signals perceived by differnt RLKs leading to a restructuring of the microbiota; this model is supported by mutants of the FERONIA (FER) RLK that show altered microbiota composition. Activated RLKs can lead to altered calcium fluxes and ROS production as well as transcriptomic changes, any and all of which can contribute to altered microbiota. (Summary by Mary Williams @PlantTeaching.bsky.social) Cell Host Microbe 10.1016/j.chom.2025.07.012
Discovery and engineering of synthetic RLKs
 Receptor-like kinases (RLKs) are abundant genes that have diversified and expanded throughout plant evolution, with most plants having hundreds or more RLKs. In this exciting new paper, Ngou et al. decided to deconstruct the recognition sites of these proteins in order to be able to make predictions about their ligands. Focusing on the LRR-RLK-XII family that encodes the familiar RLK FLS2, they clustered the sequences of the extracellular recognition domains from 13,185 genes from 350 plant genomes, and identified 210 subgroups with distinct binding domains likely to recognize different ligands, and characterized representatives for their ability to recognize live or boiled bacterial extracts in vivo. They then focused on a candidate that recognized extracts from multiple bacteria, and worked out that it recognizes a peptide csp15 from cold-shock proteins; they designated this protein SCORE (selective cold shock protein receptor). They dissected the LRR domain and identified regions that recognize different cold-shock proteins. Next they looked at the interaction between 21 SCORE orthologues versus 104 diverse csp15 peptides to map the LRR amino acids responsible for SCORE specificity. With this knowledge in hand, they designed a synthetic SCORE that recognizes a wide range of bacterial cold-shock proteins, raising the possibility of introducing broad-spectrum resistance to crops. This is truly a “wow” paper representing an impressive amount of work and valuable findings. You can also read the very helpful illustrated summary on the first author’s bluesky account here https://bsky.app/profile/brunongou.bsky.social/post/3lkmktd3ezs2g. (Summary by Mary Williams @PlantTeaching.bsky.social) Science https://10.1126/science.adx2508.
Receptor-like kinases (RLKs) are abundant genes that have diversified and expanded throughout plant evolution, with most plants having hundreds or more RLKs. In this exciting new paper, Ngou et al. decided to deconstruct the recognition sites of these proteins in order to be able to make predictions about their ligands. Focusing on the LRR-RLK-XII family that encodes the familiar RLK FLS2, they clustered the sequences of the extracellular recognition domains from 13,185 genes from 350 plant genomes, and identified 210 subgroups with distinct binding domains likely to recognize different ligands, and characterized representatives for their ability to recognize live or boiled bacterial extracts in vivo. They then focused on a candidate that recognized extracts from multiple bacteria, and worked out that it recognizes a peptide csp15 from cold-shock proteins; they designated this protein SCORE (selective cold shock protein receptor). They dissected the LRR domain and identified regions that recognize different cold-shock proteins. Next they looked at the interaction between 21 SCORE orthologues versus 104 diverse csp15 peptides to map the LRR amino acids responsible for SCORE specificity. With this knowledge in hand, they designed a synthetic SCORE that recognizes a wide range of bacterial cold-shock proteins, raising the possibility of introducing broad-spectrum resistance to crops. This is truly a “wow” paper representing an impressive amount of work and valuable findings. You can also read the very helpful illustrated summary on the first author’s bluesky account here https://bsky.app/profile/brunongou.bsky.social/post/3lkmktd3ezs2g. (Summary by Mary Williams @PlantTeaching.bsky.social) Science https://10.1126/science.adx2508.
Identification of a plastid-localized amino acid transporter
 Plastids are more than photosynthetic compartments, they also are responsible for the biosynthesis of nine proteinogenic amino acids (which explains why animals, lacking plastids, must obtain these amino acids through their diet). In a new paper, Kuhnert et al. have identified a critical amino-acid transporter on the plastid inner envelope membrane that is necessary for their export to the cytosol. The authors set out on a targeted search for this transporter by searching for genes encoding transmembrane domain proteins and plastidial target sequences that are co-expressed with amino acid biosynthetic genes. One of the candidates had previously been characterized as an Arabidopsis gene, RETICULATA1, identified as a loss-of-function mutant. The name, which means “network”, refers to the fact that the mutant has small pale-green mesophyll cells adjacent to the leaf vasculature, making the network of the green veins more prominent. Previously, this phenotype had been postulated to be due to a deficiency of essential metabolites. In the new paper, the authors noted lower levels of key amino acids in the mutants. When expressed in yeast, the RE1 protein was shown to transport basic amino acids. As the authors conclude, “These results establish RE1 as the first identified plastidial basic amino acid carrier in Arabidopsis.” (Summary by Mary Williams @PlantTeaching.bsky.social) Nature Plants 10.1038/s41477-025-02080-z
Plastids are more than photosynthetic compartments, they also are responsible for the biosynthesis of nine proteinogenic amino acids (which explains why animals, lacking plastids, must obtain these amino acids through their diet). In a new paper, Kuhnert et al. have identified a critical amino-acid transporter on the plastid inner envelope membrane that is necessary for their export to the cytosol. The authors set out on a targeted search for this transporter by searching for genes encoding transmembrane domain proteins and plastidial target sequences that are co-expressed with amino acid biosynthetic genes. One of the candidates had previously been characterized as an Arabidopsis gene, RETICULATA1, identified as a loss-of-function mutant. The name, which means “network”, refers to the fact that the mutant has small pale-green mesophyll cells adjacent to the leaf vasculature, making the network of the green veins more prominent. Previously, this phenotype had been postulated to be due to a deficiency of essential metabolites. In the new paper, the authors noted lower levels of key amino acids in the mutants. When expressed in yeast, the RE1 protein was shown to transport basic amino acids. As the authors conclude, “These results establish RE1 as the first identified plastidial basic amino acid carrier in Arabidopsis.” (Summary by Mary Williams @PlantTeaching.bsky.social) Nature Plants 10.1038/s41477-025-02080-z
Under pressure: Interdisciplinary research into the mechanics of the cell wall
 The architecture of the plant cell wall is very intricate: strings of cellulose microfibrils provide structure while allowing the cell to expand and grow. This growth is primarily driven by turgor pressure, which applies pressure and allows for expansion in all directions. While the general mechanism of growth is well established, the precise mechanical transitions that allow for extreme wall deformation without failure are not fully understood. In this recent publication by Chen et al., the authors investigated the mechanical properties of the epidermal cell wall of Arabidopsis thaliana. Using a combination of experiments and modeling, they characterized how the cell wall responds to strain at different developmental stages. They determined that the cell wall behaves like a fibrous network and that the connections between fibers explains the ability for the cell wall to stretch while maintaining structural integrity. They also found that the microfibrils exhibit non-linear stiffening, which essentially means that as growth slows, stiffness increases indicating that cells can tune their mechanical properties during development. What’s interesting about these findings is that similar behavior is observed in other biological materials like mycelium and mammalian skin. This paper blends engineering theory with plant biology, highlighting the incredible utility of interdisciplinary approaches. (Summary by Xavier Ozowara [email protected]) Nature Comms. 10.1038/s41467-025-62844-1
The architecture of the plant cell wall is very intricate: strings of cellulose microfibrils provide structure while allowing the cell to expand and grow. This growth is primarily driven by turgor pressure, which applies pressure and allows for expansion in all directions. While the general mechanism of growth is well established, the precise mechanical transitions that allow for extreme wall deformation without failure are not fully understood. In this recent publication by Chen et al., the authors investigated the mechanical properties of the epidermal cell wall of Arabidopsis thaliana. Using a combination of experiments and modeling, they characterized how the cell wall responds to strain at different developmental stages. They determined that the cell wall behaves like a fibrous network and that the connections between fibers explains the ability for the cell wall to stretch while maintaining structural integrity. They also found that the microfibrils exhibit non-linear stiffening, which essentially means that as growth slows, stiffness increases indicating that cells can tune their mechanical properties during development. What’s interesting about these findings is that similar behavior is observed in other biological materials like mycelium and mammalian skin. This paper blends engineering theory with plant biology, highlighting the incredible utility of interdisciplinary approaches. (Summary by Xavier Ozowara [email protected]) Nature Comms. 10.1038/s41467-025-62844-1
Senescence to nutrient cycling: Top-down effects of climate change
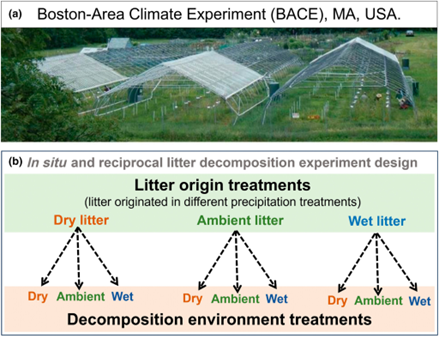 Precipitation plays an important role in shaping leaf tissue composition, and as climate change progresses, we’ll likely see dramatic shifts in precipitation regimes. Changes in leaf tissue composition due to increased rainfall or drought may have cascading effects on soil microbe communities and ultimately nutrient cycling during senescence. In this long term study, Zhang et al. explored the direct and indirect effects of climate change on leaf chemistry and soil nutrients. The authors grew Quercus rubra and Quercus velutina under dry, ambient, or wet conditions. They followed these treatments by examining the rate of decomposition of these different leaves under dry, ambient, or wet conditions. As expected, leaf litter from drought treatments contained higher concentrations of lignin and alkyl compounds (e.g., cutin, waxes, and lipids), which are synthesized in response to drought stress. Decomposition rates were variable, but they did observe faster decay with dry-grown leaves of Q. rubra under dry decomposition conditions, highlighting that leaf decay may be species specific. Lastly, they found downstream effects of dry decay conditions, with increased ergosterol in soil, which is an indicator of elevated fungal growth. While many studies focus on the effects of climate on plant structures or the soil microbiome, this paper provides a novel insight into the top-down effects of climate change in an ecosystem. (Summary by Xavier Ozowara [email protected]). New Phytol. 10.1111/nph.70449
Precipitation plays an important role in shaping leaf tissue composition, and as climate change progresses, we’ll likely see dramatic shifts in precipitation regimes. Changes in leaf tissue composition due to increased rainfall or drought may have cascading effects on soil microbe communities and ultimately nutrient cycling during senescence. In this long term study, Zhang et al. explored the direct and indirect effects of climate change on leaf chemistry and soil nutrients. The authors grew Quercus rubra and Quercus velutina under dry, ambient, or wet conditions. They followed these treatments by examining the rate of decomposition of these different leaves under dry, ambient, or wet conditions. As expected, leaf litter from drought treatments contained higher concentrations of lignin and alkyl compounds (e.g., cutin, waxes, and lipids), which are synthesized in response to drought stress. Decomposition rates were variable, but they did observe faster decay with dry-grown leaves of Q. rubra under dry decomposition conditions, highlighting that leaf decay may be species specific. Lastly, they found downstream effects of dry decay conditions, with increased ergosterol in soil, which is an indicator of elevated fungal growth. While many studies focus on the effects of climate on plant structures or the soil microbiome, this paper provides a novel insight into the top-down effects of climate change in an ecosystem. (Summary by Xavier Ozowara [email protected]). New Phytol. 10.1111/nph.70449
A wheat immune receptor that breaks the mold: The octameric WAI3 resistosome
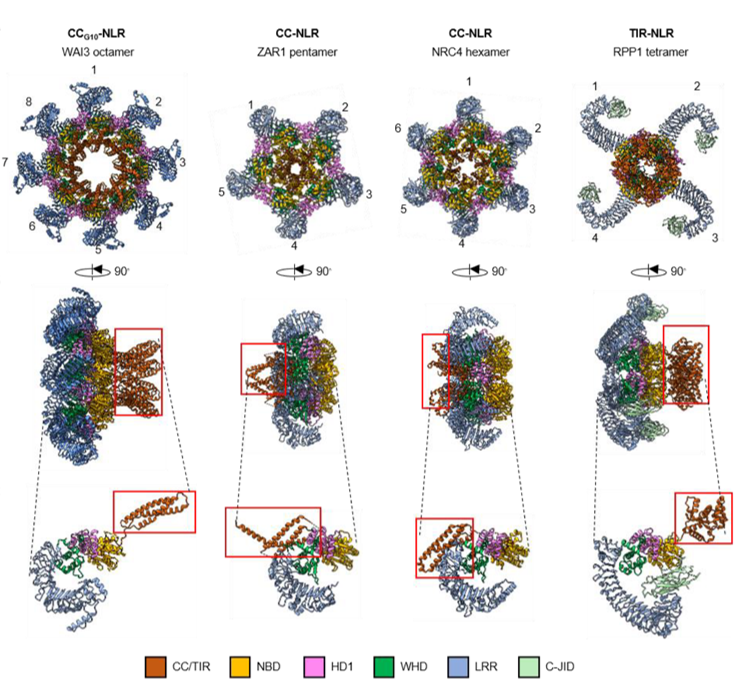 Plants may not have white blood cells, but they are far from defenseless. Instead, they rely on specialized immune receptors called NLRs (nucleotide-binding leucine-rich repeat receptors) to recognize intracellular invaders and trigger strong defense responses. These receptors are molecular shape-shifters, assembling into large multi-protein complexes known as resistosomes. For example, the ZAR1 resistosome forms a pentamer, NRC4 assembles as a hexamer, and RPP1 as a tetramer. In a recent study, Guo and colleagues uncovered an unusual case in wheat through a forward genetic screen. They identified an EMS mutant in the gene Wheat Autoimmunity 3 (WAI3), which shows constant activation of plant immunity. WAI3 belongs to a unique sub-clade of G10-type coiled-coil NLRs, distinct from other CC-NLRs. When expressed in Nicotiana benthamiana, a truncated (inactivated) version of WAI3 unexpectedly formed an octameric resistosome—the largest oligomeric assembly reported for plant NLRs so far. Intriguingly, WAI3 lacks the EDVID domain, a structural element that stabilizes domain interactions in other resistosomes. This absence results in a distinct domain arrangement and LRR topology. While WAI3 still exhibits calcium channel properties like ZAR1, the significance of its unusual architecture remains an exciting open question for future research. (Summary by Ching Chan @ntnuchanlab) bioRxiv 10.1101/2025.08.26.672026
Plants may not have white blood cells, but they are far from defenseless. Instead, they rely on specialized immune receptors called NLRs (nucleotide-binding leucine-rich repeat receptors) to recognize intracellular invaders and trigger strong defense responses. These receptors are molecular shape-shifters, assembling into large multi-protein complexes known as resistosomes. For example, the ZAR1 resistosome forms a pentamer, NRC4 assembles as a hexamer, and RPP1 as a tetramer. In a recent study, Guo and colleagues uncovered an unusual case in wheat through a forward genetic screen. They identified an EMS mutant in the gene Wheat Autoimmunity 3 (WAI3), which shows constant activation of plant immunity. WAI3 belongs to a unique sub-clade of G10-type coiled-coil NLRs, distinct from other CC-NLRs. When expressed in Nicotiana benthamiana, a truncated (inactivated) version of WAI3 unexpectedly formed an octameric resistosome—the largest oligomeric assembly reported for plant NLRs so far. Intriguingly, WAI3 lacks the EDVID domain, a structural element that stabilizes domain interactions in other resistosomes. This absence results in a distinct domain arrangement and LRR topology. While WAI3 still exhibits calcium channel properties like ZAR1, the significance of its unusual architecture remains an exciting open question for future research. (Summary by Ching Chan @ntnuchanlab) bioRxiv 10.1101/2025.08.26.672026
Signals in the air and soil: the hidden communication circuit that shapes plant resilience
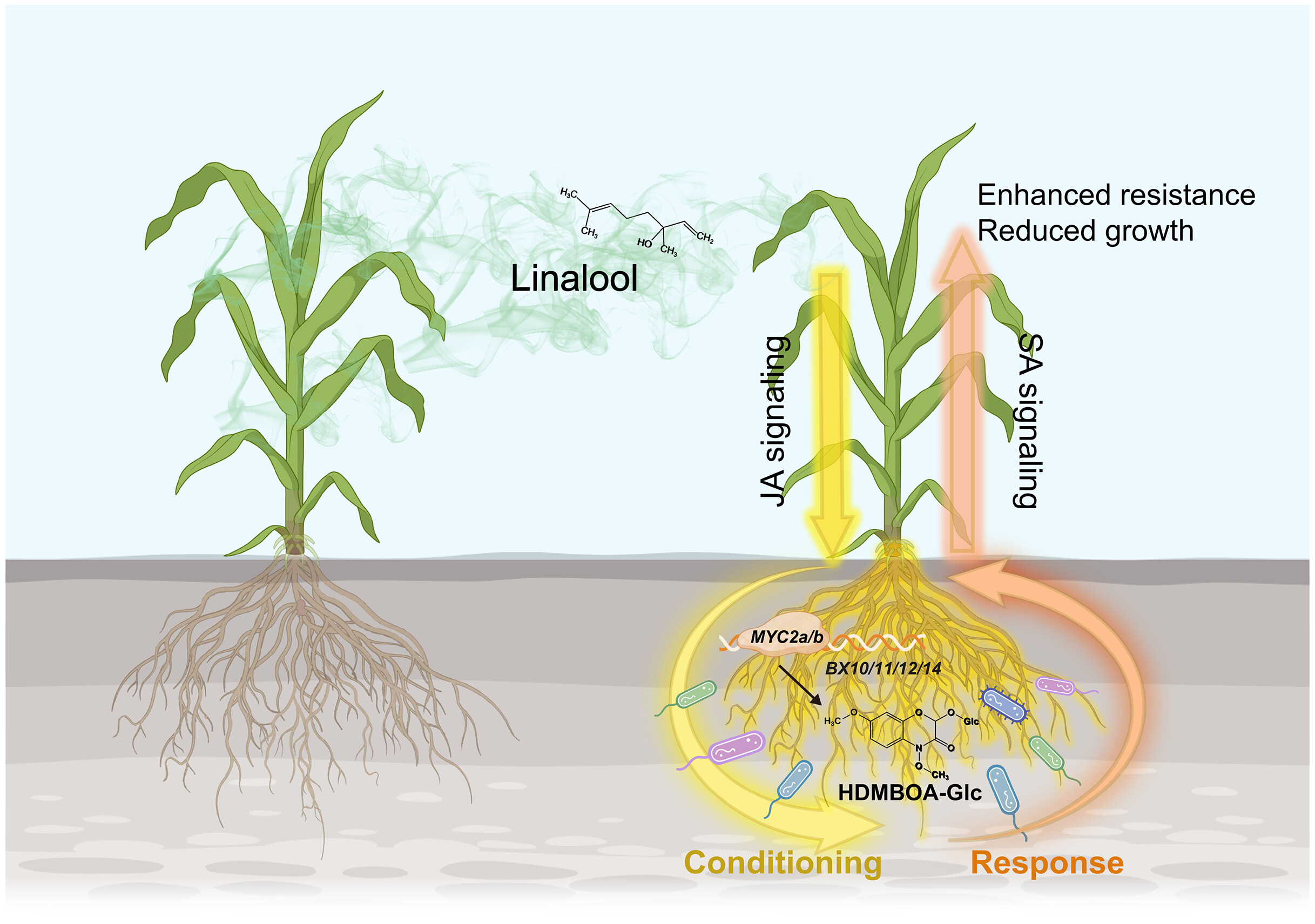 When plants grow shoulder to shoulder in crowded fields, they don’t just compete for space and sunlight—they also “talk” to one another. This underground and airborne chatter, carried by leaf volatiles and root exudates, helps plants sense nearby neighbors and prepare for threats. But how exactly does this communication unfold? Guo and colleagues set out to untangle this mystery using maize. They first grew plants at different densities, each in its own pot, then used the conditioned soil to raise new plants. Remarkably, seedlings grown in soil from high-density environments grew smaller but were far tougher against herbivores, fungi, and viruses. Something in the soil was clearly transmitting a memory of crowding. Digging deeper, the team traced the signal back to a leaf volatile: linalool. This compound activates jasmonate signaling in neighboring plants, triggering the roots to release benzoxazinoids. These chemicals reshape the rhizosphere microbiome, which in turn bolsters plant immunity – at the cost of growth. The study uncovers a remarkable feedback circuit, showing how maize reprograms itself in crowded settings by channeling energy from growth into defense. (Summary by Ching Chan @ntnuchanlab) Science 10.1126/science.adv6675
When plants grow shoulder to shoulder in crowded fields, they don’t just compete for space and sunlight—they also “talk” to one another. This underground and airborne chatter, carried by leaf volatiles and root exudates, helps plants sense nearby neighbors and prepare for threats. But how exactly does this communication unfold? Guo and colleagues set out to untangle this mystery using maize. They first grew plants at different densities, each in its own pot, then used the conditioned soil to raise new plants. Remarkably, seedlings grown in soil from high-density environments grew smaller but were far tougher against herbivores, fungi, and viruses. Something in the soil was clearly transmitting a memory of crowding. Digging deeper, the team traced the signal back to a leaf volatile: linalool. This compound activates jasmonate signaling in neighboring plants, triggering the roots to release benzoxazinoids. These chemicals reshape the rhizosphere microbiome, which in turn bolsters plant immunity – at the cost of growth. The study uncovers a remarkable feedback circuit, showing how maize reprograms itself in crowded settings by channeling energy from growth into defense. (Summary by Ching Chan @ntnuchanlab) Science 10.1126/science.adv6675
More than a trade-off: how growth inhibition protects plant genome integrity
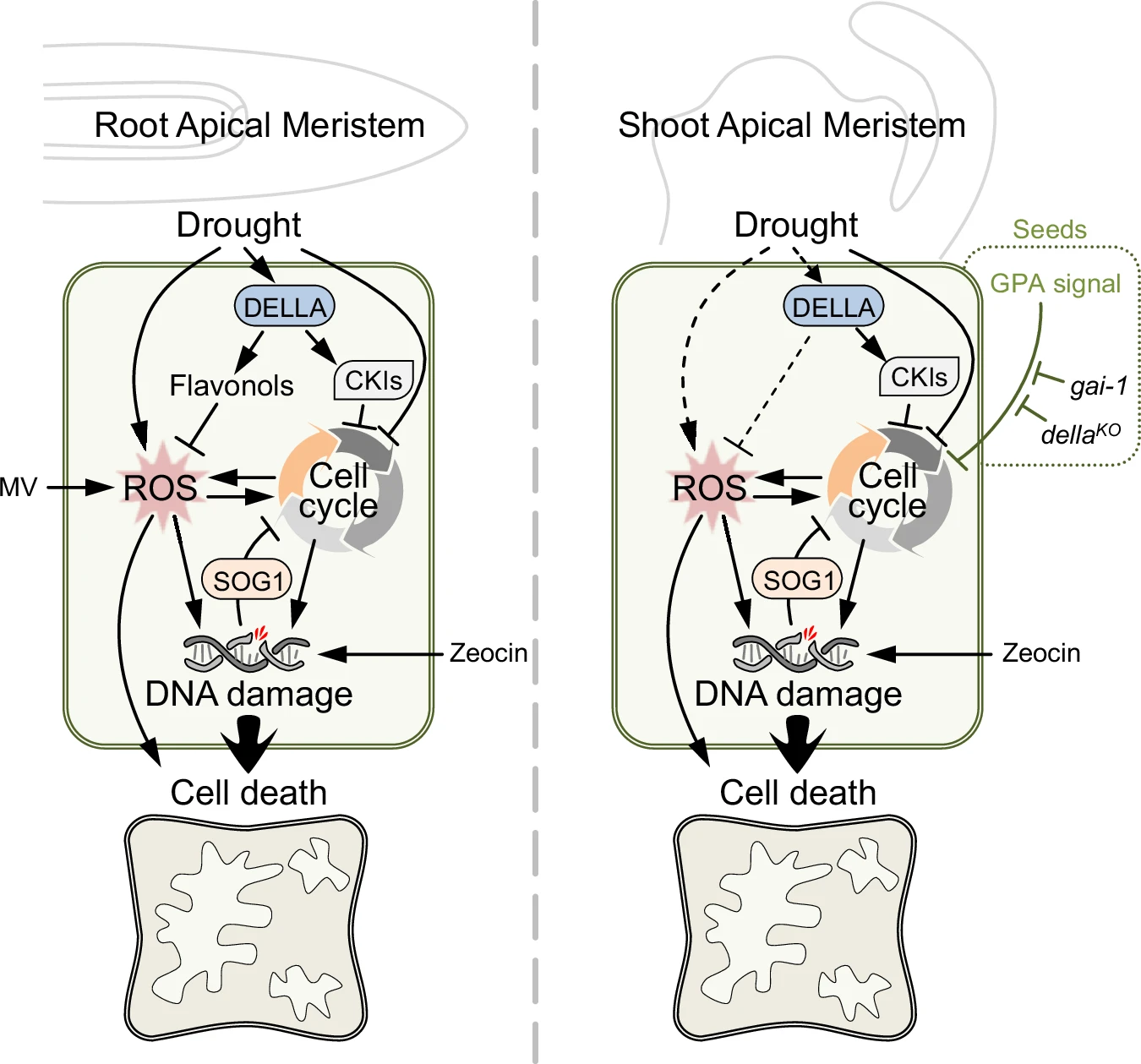 When plants face tough times, like an invading pathogen or a sudden drought, they often make a difficult choice: pause growth to focus on survival. This phenomenon, called the growth–defense trade-off, has long been thought to result from limited resources – plants simply can’t do everything at once. But recent discoveries suggest the story is more complex. Sometimes, plants can maintain resilience without giving up growth, depending on how key signaling pathways are tuned. This raises a deeper question: is slowing growth just about saving resources, or does it serve another hidden purpose? To explore this, Serrano-Mislata and colleagues studied the role of DELLA proteins, well-known growth repressors, during dehydration stress. They found that disabling DELLA’s targets – the CDK inhibitors (CKIs) in shoots or roots – allowed plants to keep growing. Yet, this came at a price: the plants lost more water and needed to boost antioxidant levels and accumulate flavonols to survive. Therefore, the authors proposed that growth cessation itself might provide protection. By assessing the level of DNA damage as a result of cell cycle arrest, della mutants exhibited elevated ROS, DNA damage, and cell death in the apical meristems. The study reveals growth arrest not as a cost of stress or weakness, but as a built-in safeguard for genome integrity and survival. (Summary by Ching Chan @ntnuchanlab) Nature Communications 10.1038/s41467-025-60733-1
When plants face tough times, like an invading pathogen or a sudden drought, they often make a difficult choice: pause growth to focus on survival. This phenomenon, called the growth–defense trade-off, has long been thought to result from limited resources – plants simply can’t do everything at once. But recent discoveries suggest the story is more complex. Sometimes, plants can maintain resilience without giving up growth, depending on how key signaling pathways are tuned. This raises a deeper question: is slowing growth just about saving resources, or does it serve another hidden purpose? To explore this, Serrano-Mislata and colleagues studied the role of DELLA proteins, well-known growth repressors, during dehydration stress. They found that disabling DELLA’s targets – the CDK inhibitors (CKIs) in shoots or roots – allowed plants to keep growing. Yet, this came at a price: the plants lost more water and needed to boost antioxidant levels and accumulate flavonols to survive. Therefore, the authors proposed that growth cessation itself might provide protection. By assessing the level of DNA damage as a result of cell cycle arrest, della mutants exhibited elevated ROS, DNA damage, and cell death in the apical meristems. The study reveals growth arrest not as a cost of stress or weakness, but as a built-in safeguard for genome integrity and survival. (Summary by Ching Chan @ntnuchanlab) Nature Communications 10.1038/s41467-025-60733-1