Paving the way for C4 evolution: study of C3-C4 intermediate species in grasses
C4 photosynthesis originated as an adaptation to low atmospheric CO2 concentrations 30-35 million years ago (Sage, 2016). Although accounting for only 3% of the vascular plants on earth, C4 plants contribute 25% of terrestrial photosynthesis (Westhoff and Gowik, 2010), thus providing a highly desirable system to increase carbon fixation efficiency and enhance crop yield. The transition from C3 to C4 photosynthesis has occurred more than 62 times independently in a complex process with a combination of anatomical, biochemical and physiological changes in leaves. In spite of their multiple origins, C4 plants are only present in certain angiosperm families, including about 5000 grasses, 1300 sedges, and 1800 dicot species, indicating developmental and anatomical constraints for C4 evolution (Sage, 2016).
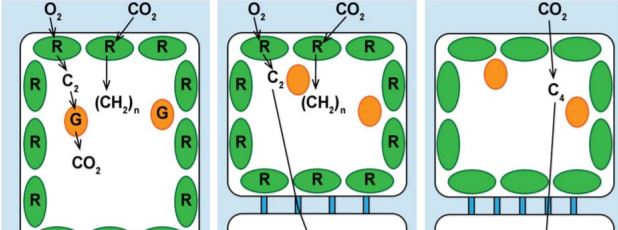
In C4 photosynthesis, CO2 is initially taken up by mesophyll cells and converted into a four-carbon (C4) compound by phosphoenolpyruvate carboxylase (PEPC), which has high affinity for CO2. Then, the C4 compound is transported to the chloroplasts of the bundle sheath cells where it releases CO2, resulting in increased CO2 concentration and enhanced carboxylase activity of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), the rate limiting enzyme of photosynthesis. The mesophyll and bundle sheath are arranged in a “wreath-like” shape surrounding the vascular bundle, a structure referred to as the Kranz anatomy. In grasses and sedges, there are two layers of sheath cells, an inner bundle sheath (also called mestome sheath) and an outer bundle sheath. One or both types of sheath cells may be co-opted for C4 function, although one of them is often lost during C4 evolution (Lundgren et al., 2014).
A few anatomical and cellular adaptations are required for efficient C4 physiology: (a) compartmentation of CO2 assimilation and fixation by increasing PEPC activity in the mesophyll cells and limiting Rubisco only to bundle sheath chloroplasts, (b) increased size and number of chloroplasts in the bundle sheath, (c) short distance between mesophyll and bundle sheath cells by increasing vein density and/or enlarging bundle sheath and mesophyll cell size, and (d) enrichment of plasmodesmata between mesophyll and bundle sheath cells to facilitate metabolic exchange (Fig. 1) (Lundgren et al., 2014; Sage, 2016). If the evolution of these traits is stepwise or progressive, we may expect to observe species with intermediate traits between C3 and C4 during evolutionary history. Indeed, C2 photosynthesis that utilizes glycine (two-carbon compound, C2) as a shuttle to concentrate CO2 is proposed as an intermediate state during C4 evolution. It is worth noting that although grasses and sedges dominate C4 species, only less than 20% of known intermediate species are monocots, hindering our understanding of C4 evolution within monocots (Sage et al., 2014). In this issue of Plant Physiology, Khoshravesh et al. (2019) compared various anatomical, ultrastructural and biochemical traits in nine species of the subtribe Neurachninae of the grass family, and identified new C3-C4 intermediate species.
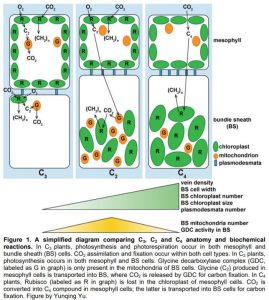 C2 photosynthesis differs from C4 in biochemical reactions in that C2 plants concentrate CO2 by limiting the photorespiratory reactions in the mitochondria of bundle sheath cells. Glycine generated by oxygenation reaction of Rubisco in mesophyll cells is transported to the bundle sheath cells and then converted to serine and CO2 by the mitochondrial glycine decarboxylase complex (GDC) (Fig. 1). Therefore, a major feature of C2 anatomy is increased abundance of mitochondria with high GDC activity, and aggregation of mitochondria and chloroplasts in the inner side of bundle sheath cells to minimize gas exchange with mesophyll cells (Fig. 1). The close association between mitochondria and chloroplasts also facilitates diffusion of photorespired CO2 into the chloroplasts, and thus concentrates CO2 surrounding Rubisco (Sage et al., 2014). C2 anatomy exhibits several C4-like features (although less pronounced than C4), including prominent bundle sheath cells, increased vein density, more abundant and asymmetrically distributed chloroplasts in the bundle sheath, and higher plasmodesmatal frequency (Fig. 1) (Sage et al., 2014; Khoshravesh et al., 2019).
C2 photosynthesis differs from C4 in biochemical reactions in that C2 plants concentrate CO2 by limiting the photorespiratory reactions in the mitochondria of bundle sheath cells. Glycine generated by oxygenation reaction of Rubisco in mesophyll cells is transported to the bundle sheath cells and then converted to serine and CO2 by the mitochondrial glycine decarboxylase complex (GDC) (Fig. 1). Therefore, a major feature of C2 anatomy is increased abundance of mitochondria with high GDC activity, and aggregation of mitochondria and chloroplasts in the inner side of bundle sheath cells to minimize gas exchange with mesophyll cells (Fig. 1). The close association between mitochondria and chloroplasts also facilitates diffusion of photorespired CO2 into the chloroplasts, and thus concentrates CO2 surrounding Rubisco (Sage et al., 2014). C2 anatomy exhibits several C4-like features (although less pronounced than C4), including prominent bundle sheath cells, increased vein density, more abundant and asymmetrically distributed chloroplasts in the bundle sheath, and higher plasmodesmatal frequency (Fig. 1) (Sage et al., 2014; Khoshravesh et al., 2019).
In addition to typical C2 photosynthesis, other C3-C4 intermediate species with different intermediate traits may shed light on C4 evolution. Khoshravesh et al. (2019) focused on subtribe Neurachninae, which contains a well-known C2 species (Neurachne minor), two C4 species (N. munroi and N. muelleri), and several putative C3 species (N. alopecuroidea, N. queenslandica, Thyridolepis mitchelliana, N. tenuifolia, N. annularis and N. lanigera). The Neurachne species possess two layers of bundle sheath, an inner suberized mestome sheath where carbon fixation occurs (functional bundle sheath), and an outer parenchymatous sheath that separate mesophyll cells and mestome sheath. Khoshravesh et al. (2019) measured C* (CO2 compensation point) as an indicator of photorespiratory CO2 refixation. They further comprehensively quantified traits including mestome sheath and mesophyll cell size in three dimensions, ratio of different cell types, mitochondrial and chloroplast number and size, vein spacing, plasmodesmatal frequency, and the abundance of glycine decarboxylase subunit P (GLDP) and Rubisco large subunit by immunolabeling. These analyses revealed a linear relationship between the above traits and C*, indicating the importance of these anatomical and biochemical characters contributing to photosynthetic efficiency.
Furthermore, among the putative C3 species, N. lanigera and N. annularis exhibited various degrees of intermediate traits between typical C3 and C2. For example, compared to typical C3 species, N. lanigera has significantly shorter and wider mestome sheath cells, increased number of chloroplasts and mitochondria in mestome sheath, greater GLDP labeling and enriched plasmodesmata. Consistently, C* of N. lanigera is significantly lower than typical C3 species particularly at higher temperature (35°C), although it is still higher than C* of the C2 species N. minor. In contrast, N. annularis functions as a C3 as its C* is similar to that of other C3 species. However, it shows subtle C2-like characters, including slightly wider mestome sheath cells and greater GLDP abundance per mestome sheath chloroplast, indicating its early stage towards C2 evolution (Khoshravesh et al., 2019).
Unlike previous phylogenetic analyses showing that C3-C4 species indeed represent an intermediate state towards C4 evolution (Schlüter and Weber, 2016), the progressive C4 traits of the Neurachne species do not reflect the phylogenetic relationships at the species level. In other words, an ancestral state with C3-C4 intermediacy may lead to the evolution of C4, C2 or even C3 species, suggesting that C3 and C3-C4 intermediates may represent partial reversals, having evolved from a C4-like ancestor. Analyses of more tribes or clades with multiple C3-C4 species may further advance our understanding of the direction of C3/C4 evolution.
Yunqing Yu, Assistant Features Editor
Donald Danforth Plant Science Center
Saint Louis, MO, USA
References
Khoshravesh R, Stata M, Busch FA, Saladie M, Castelli JM, Dakin N, Hattersley PW, Macfarlane TD, Sage RF, Ludwig M (2019) The evolutionary origin of C4 photosynthesis in the grass subtribe Neurachninae. Plant Physiology: pp. 00925.02019 https://doi.org/10.1104/pp.19.00925
Lundgren MR, Osborne CP, Christin P-A (2014) Deconstructing Kranz anatomy to understand C4 evolution. Journal of Experimental Botany 65: 3357-3369
Sage RF (2016) A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, and Hall of Fame. Journal of Experimental Botany 68: e11-e28
Sage RF, Khoshravesh R, Sage TL (2014) From proto-Kranz to C4 Kranz: building the bridge to C4 photosynthesis. Journal of Experimental Botany 65: 3341-3356
Schlüter U, Weber AP (2016) The road to C4 photosynthesis: evolution of a complex trait via intermediary states. Plant and Cell Physiology 57: 881-889
Westhoff P, Gowik U (2010) Evolution of C4 photosynthesis—looking for the master switch. Plant Physiology 154: 598-601