Letting Sleeping DOGs Lie: Regulation of DOG1 during Seed Dormancy
The auspicious timing of seasonal germination is crucial to plant fitness and survival. Consequently, seed dormancy is tightly regulated by both developmental and environmental cues, which are integrated via crosstalk between several phytohormone signaling pathways including gibberellins, abscisic acid (ABA), and ethylene. Since favorable environmental conditions, notably permissive temperatures and photoperiods, are crucial for the developmental switch from dormancy to germination, Arabidopsis thaliana accessions originating from diverse geographical locations exhibit significant phenotypic variation in dormancy duration. Quantitative trait loci analysis of this natural variation identified DELAY OF GERMINATION1 (DOG1) as a major regulator of dormancy, with DOG1 protein levels accumulating during seed maturation and correlating with the depth of dormancy (Bentsink et al., 2006).
Despite being studied extensively over the last decade, little is known about the precise molecular mechanism underlying the transcriptional regulation of DOG1. Now, two new studies from the labs of Yongxiu Liu (Li et al., 2019) and Peter Eastmond (Bryant et al., 2019) provide mechanistic insights into how ethylene signaling and low temperatures, respectively, control seed dormancy via DOG1 regulation.
Following on from their previous research (Mendes et al., 2013), Bryant and co-workers performed functional analysis on gain- and loss-of-function mutants of both bZIP67 and DOG1 to establish that bZIP67 is required for DOG1 expression. Indeed, ChIP-qPCR and transactivation assays revealed that bZIP67 binds directly to the DOG1 promoter and that this interaction also requires the expression of the four known master regulators of seed maturation, namely LEC1, ABI3, FUS3, and LEC2. Crucially, within this DOG1 regulon, bZIP67 modulates DOG1 expression in response to low environmental temperatures via a posttranscriptional increase in bZIP67 protein abundance, ultimately enhancing seed dormancy status.
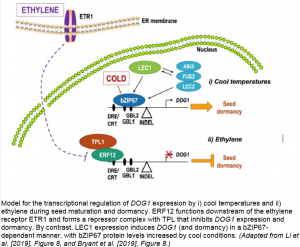 Whereas cool conditions promote seed dormancy, exogenous ethylene application induces germination (Linkies and Leubner-Metzger, 2012). Through the detailed characterization of an Arabidopsis mutant with significantly reduced dormancy (later identified as a loss-of-function mutant of the ethylene receptor, ETR1), Li and colleagues demonstrated that DOG1 functions downstream of both ETR1 and the ethylene response factor, ERF12. In fact, ERF12 binds directly to the DOG1 promoter, recruiting the co-repressor TOPLESS in the process and inhibiting DOG1 expression.
Whereas cool conditions promote seed dormancy, exogenous ethylene application induces germination (Linkies and Leubner-Metzger, 2012). Through the detailed characterization of an Arabidopsis mutant with significantly reduced dormancy (later identified as a loss-of-function mutant of the ethylene receptor, ETR1), Li and colleagues demonstrated that DOG1 functions downstream of both ETR1 and the ethylene response factor, ERF12. In fact, ERF12 binds directly to the DOG1 promoter, recruiting the co-repressor TOPLESS in the process and inhibiting DOG1 expression.
In successfully establishing bZIP67 and ERF12 as direct regulators of DOG1, these two studies also identified their corresponding binding sites, namely two G box-like (GBL) motifs and a DRE/CRT element, respectively. Further examination of the DOG1 promoter sequence reveals that these cis-elements are actually in very close proximity to each other, located at -456 bp (GBL1), -500 bp (GBL2), and -646 bp (DRE/CRT) upstream of the DOG1 start codon. This region of the promoter may therefore represent an epicenter for binding of transcription factor complexes involved in combinatorial regulation of DOG1. Given that the DRE/CRT regulon is associated with cold responses and is ABA-inducible (Knight et al., 2004), it is also conceivable that ERF12 may play a wider role in DOG1 regulation beyond that linked to ethylene signaling.
Immediately downstream of the GBL1 cis-element is a 285 bp sequence that is present in the DOG1 promoter of the weakly dormant Ler-0 accession but, crucially, is absent from the Cvi-0 ecotype, which exhibits strong dormancy. This INDEL was found to directly affect the ability of bZIP67 to transactivate DOG1 in vivo and may explain the observed variation in DOG1 transcript levels, and consequently dormancy, between ecotypes. Such natural allelic variation in DOG1 coupled with DOG1 expression plasticity confers substantial adaptive significance in the field, where seasonal environmental factors can vary enormously and the optimal timing of seed germination is paramount.
REFERENCES
Bentsink, L., Jowett, J., Hanhart, C.J., and Koornneef, M. (2006). Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proceedings of the National Academy of Sciences 103: 17042–17047.
Bryant, F.M., Hughes, D., Hassani-Park, K., Eastmond P.J. (2019). Basic LEUCINE ZIPPER TRANSCRIPTION FACTOR 67 transactivates DELAY OF GERMINATION 1 to establish primary seed dormancy in Arabidopsis. Plant Cell https://doi.org/10.1105/tpc.18.00892.
Knight, H., Zarka, D.G., Okamoto, H., Thomashow, M.F., and Knight, M.R. (2004). Abscisic acid induces CBF gene transcription and subsequent induction of cold-regulated genes via the CRT promoter element. Plant Physiology 135: 1710–1717.
Li, X., Chen, T., Li, Y., Wang, Z., Cao, H., Chen, F., Li, Y., Soppe, W.J.J., Li, W., Liu, Y. (2019). ETR1/RDO3 Regulates Seed Dormancy by Relieving the Inhibitory Effect of the ERF12-TPL Complex on DELAY OF GERMINATION1 Expression. Plant Cell https://doi.org/10.1105/tpc.18.00449.
Linkies, A. and Leubner-Metzger, G. (2012). Beyond gibberellins and abscisic acid: how ethylene and jasmonates control seed germination. Plant Cell Rep 31: 253–270.
Mendes, A., Kelly, A.A., van Erp, H., Shaw, E., Powers, S.J., Kurup, S., and Eastmond, P.J. (2013). bZIP67 regulates the omega-3 fatty acid content of Arabidopsis seed oil by activating fatty acid desaturase3. Plant Cell 25: 3104–3116.
