Leaving the boundary behind to make a new branch
Nicolas et al. explore the mechanisms regulating CUP-SHAPED COTYLEDON expression in axillary meristem initiation. https://doi.org/10.1093/plcell/koac269
Antoine Nicolasa,b and Patrick Laufsa
a Université Paris-Saclay, INRAE, AgroParisTech, Institut Jean-Pierre Bourgin (IJPB), 78000, Versailles, France.
b Université Paris-Saclay, 91405 Orsay, France
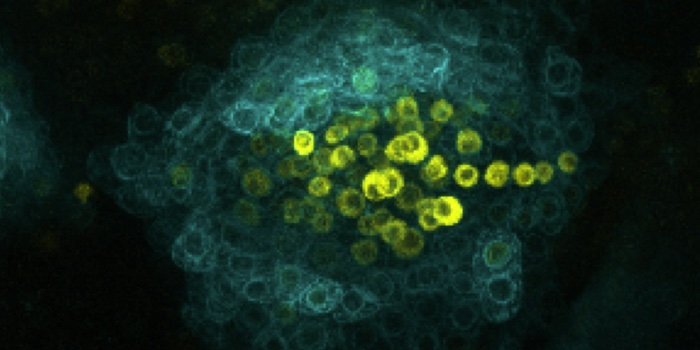
Background: Stem cells function as sources of new cells for growth or replacing injured tissues. In plants, all tissues are descended from stem cells located in their growing tips, within the meristems, which are therefore essential for forming the shoot and root systems. Some of these meristems form during embryogenesis, but others form later. For example, the axillary meristems that produce lateral shoots arise from a few cells in the boundary domain at the junction between the leaf and the stem. The CUP-SHAPED COTYLEDON (CUC) genes are major determinants of the boundary domain and are required for initiating these axillary meristems. However, little is known about the later steps that lead to well-organized axillary meristems and, in particular, to the de novo establishment of stem cells.
Questions: How are axillary meristems structured during development? How is a new stem cell pool established de novo in an axillary meristem?
Findings: We mainly focused on the establishment of cauline axillary meristems in Arabidopsis thaliana because they are poorly characterized compared to rosette axillary meristems. Cauline axillary meristems form rapidly after the floral transition and we described the dynamic modification of gene expression that occurs along with their formation. Accordingly, while the CUC genes are required for the early steps of axillary meristem initiation, they have to be cleared for meristem establishment and activation of a stem cell population. We showed that two NGATHA-LIKE transcription factors are required to repress CUC expression in the initiating axillary meristem (cauline and rosette axillary meristems) to permit de novo establishment of stem cells. Because we observed that stem cell formation is also affected in floral meristems of double mutants for these two NGATHA-LIKE transcription factors, our work reveals a mechanism operating in different types of newly formed meristems.
Next steps: One of the next steps would be to connect the role of the NGATHA-LIKE–CUC interaction during axillary meristem formation with other well-known regulators such as cytokinin signaling or HAM transcription factors. In addition, it would be interesting to test for a molecular connection between CUC transcription factors and genes controlling stem cell fate.
Reference:
Antoine Nicolas, Aude Maugarny-Calès, Bernard Adroher, Liudmila Chelysheva, Yu Li, Jasmine Burguet, Anne-Maarit Bågman, Margot E. Smit, Siobhan M. Brady, Yunhai Li, Patrick Laufs (2022) De novo stem cell establishment in meristems requires repression of organ boundary cell fate. Plant Cell. https://doi.org/10.1093/plcell/koac269







