Attract or Defend: The CYP-associated Versatility of Terpenoids
Floral scent, a blend of volatile organic compounds (VOCs), is a communication signal central to plant interactions. For example, VOCs attract pollinators when flowers are opened (Muhleman et al., 2015).
However, some plants do not cross-pollinate and depend less on VOCs. Flowers are fragile structures with fewer physical barriers than other plant tissues, which can make them targets for pathogens and florivores. In outcrossing plants, this vulnerability is compensated by successful reproduction. However, self-pollinating plants have a fitness advantage in balancing VOC release with defense.
Arabidopsis thaliana is a good model to research how self-pollinated plants reduce VOCs. Arabidopsis flowers emit a complex but weak blend of terpenoid volatiles. Flowers were shown to express 20 terpene synthases (TPSs), but the role of these genes was unclear until recently. One hypothesis was that TPS gene expression was a vestigial trait (Chen et al., 2003).
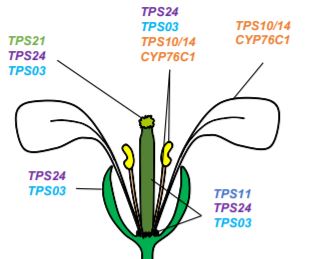
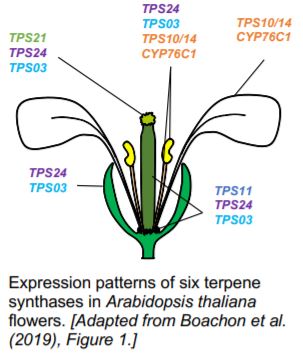 A decade later, TPS enzymes revealed their key role (Ginglinger et al., 2013). Indeed, Arabidopsis plants reuse the terpenoid pathways for defense instead of attraction. Volatile terpenes are still synthesized; however, they are further oxidized into non-volatile defensive chemicals (Boachon et al., 2015; Boachon et al., 2019). Soluble terpenes accumulate within tissues at the expense of volatiles, explaining the strong reduction of floral scent.
A decade later, TPS enzymes revealed their key role (Ginglinger et al., 2013). Indeed, Arabidopsis plants reuse the terpenoid pathways for defense instead of attraction. Volatile terpenes are still synthesized; however, they are further oxidized into non-volatile defensive chemicals (Boachon et al., 2015; Boachon et al., 2019). Soluble terpenes accumulate within tissues at the expense of volatiles, explaining the strong reduction of floral scent.
Multifunctional cytochrome P450 enzymes (CYPs) with low substrate specificity reduce the volatility of terpenes in Arabidopsis flowers. Boachon et al. (2015) showed that CYP76C1, a monoterpene oxidase, oxidizes linalool enantiomers produced by TPS10 and TPS14 into lilac compounds. In their most recent paper, Boachon et al. (2019) highlighted the role of CYP706A3 in the oxidation of VOCs. CYP706A3 clusters with TPS11 on chromosome 5 and the two genes are co-expressed. However, CYP706A3 does not oxidize only the sesquiterpenes produced by TPS11. CYP706A3 also accepts (E)-β-caryophyllene and various monoterpenes as substrates. Overall, CYP706A3 and CYP76C1 reduce the volatility of the compounds synthesized by TPS10, TPS11, TPS14, TPS21, and TPS24 (Figure). Other CYP enzymes may also contribute to these oxidations in cascade (Ginglinger et al., 2013).
Both Boachon et al. (2015) and Boachon et al. (2019) show the benefits of this strategy associated with self-pollination. Linalool attracts thrips and hoverflies. Instead, lilac compounds reduce these visits and the resulting damage by three other feeding insects (Boachon et al., 2015). Similarly, sesquiterpenes produced by TPS11 attract hoverflies while their oxidized derivatives repel them. As the expression of TPS11 and CYP706A3 start early in flower development, Boachon et al. (2019) further tested the feeding behavior of Plutella xylostella caterpillars and showed that they avoided floral buds with oxidized terpenes. Finally, microbiome analyses suggest that flowers emitting a blend of VOCs may host different bacterial communities than flowers accumulating non-volatile terpenes.
Céline Caseys, Assistant Features Editor
Department of Plant Sciences, University of California, Davis
REFERENCES
Boachon B, Junker RR, Miesch L, Bassard JE, Höfer R, Caillieaudeaux R, Seidel DE, Lesot A, Heinrich C, Ginglinger JF, Allouche L. (2015). CYP76C1 (Cytochrome P450)-mediated linalool metabolism and the formation of volatile and soluble linalool oxides in Arabidopsis flowers: a strategy for defense against floral antagonists. The Plant Cell, 27(10): 2972-90.
Boachon B, Burdloff Y, Ruan Y-X, Rojoe R, Junker RR, Vincent B, Nicolè F, Bringel F, Lesot A, Henry L, Bassard J-H, Mathieu S, Allouche L, Kaplan I, Dudareva N, Vuilleumier S, Miesch L, André F, Navrot N, Chen X-Y, Werck-Reichhart D. (2019). A Promiscuous CYP706A3 Reduces Terpene Volatile Emission from Arabidopsis Flowers, with Impacts on Florivores and Floral Microbiome. The Plant Cell DOI: https://doi.org/10.1105/tpc.19.00320.
Chen F, Tholl D, D’Auria JC, Farooq A, Pichersky E, Gershenzon J. (2003). Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers. The Plant Cell, 15(2), 481-494.
Ginglinger JF, Boachon B, Höfer R, Paetz C, Köllner TG, Miesch L, Lugan R, Baltenweck R, Mutterer J, Ullmann P, Beran F. (2013). Gene coexpression analysis reveals complex metabolism of the monoterpene alcohol linalool in Arabidopsis flowers. The Plant Cell, 25(11):4640-57.
Muhlemann JK, Klempien A, Dudareva N. (2014) Floral volatiles: from biosynthesis to function. Plant, cell & environment, 37(8):1936-49.

