Plant Science Research Weekly: February 24, 2023
Review: Photorespiration is the solution, not the problem
 Rubisco (ribulose 1,5-bisphosphate carboxylase/oxygenase) is one of a kind, simultaneously recognized as one of the most abundant and important enzymes, and also widely characterized as flawed because it uses both O2 and CO2 as substrates, leading to both carboxylation and oxygenation of ribulose bisphosphate. When CO2 is used as a substrate, the carbon is fixed and converted to sugars, but when O2 is the substrate the product, 2-phosphoglycolate, must be removed through the photorespiratory pathway (consuming ATP) to avoid inhibiting rubisco. Several strategies to improve photosynthesis have bene explored, many of which suppress photorespiration, but, as discussed in a Humboldt Revew by Bauwe and further highlighted in a commentary by Brocano et al., photorespiration is not itself the problem but rather the solution. The Bauwe review is encyclopedic and essential to those who work in the field, whereas the Brocano et al. article beautifully distills the essences for the rest of us. (Summary by Mary Williams @PlantTeaching) J. Plant Physiol. 10.1016/j.jplph.2022.153899 and 10.1016/j.jplph.2023.153928
Rubisco (ribulose 1,5-bisphosphate carboxylase/oxygenase) is one of a kind, simultaneously recognized as one of the most abundant and important enzymes, and also widely characterized as flawed because it uses both O2 and CO2 as substrates, leading to both carboxylation and oxygenation of ribulose bisphosphate. When CO2 is used as a substrate, the carbon is fixed and converted to sugars, but when O2 is the substrate the product, 2-phosphoglycolate, must be removed through the photorespiratory pathway (consuming ATP) to avoid inhibiting rubisco. Several strategies to improve photosynthesis have bene explored, many of which suppress photorespiration, but, as discussed in a Humboldt Revew by Bauwe and further highlighted in a commentary by Brocano et al., photorespiration is not itself the problem but rather the solution. The Bauwe review is encyclopedic and essential to those who work in the field, whereas the Brocano et al. article beautifully distills the essences for the rest of us. (Summary by Mary Williams @PlantTeaching) J. Plant Physiol. 10.1016/j.jplph.2022.153899 and 10.1016/j.jplph.2023.153928
Review: One plant’s poison
 Plants synthesize interesting chemicals that attract, deter, amuse, and harm their predators. Some of the most harmful to humans have been selectively eliminated through the process of domestication, but others render potential food sources inedible. This review by Liu et al. discusses four approaches to increasing harvest index (product per plant) by selectively removing phytotoxins from otherwise edible tissues. As an example, antinutritional glucosinolates (GSLs) in canola seeds make the residue after extracting the oil inedible. Similarly, although cotton seeds have a lot of protein, they also contain phytotoxic gossypol, and cassava leaves contain cyanogenic glycosides. For these and other phytotoxins, several strategies have been explored to eliminate them from tissues that would otherwise be good food sources. These strategies include interfering with their biosynthetic enzymes or the genes that encode them, disrupting their transport or storage, or converting them to non-toxic forms. As the authors observe, detoxified plants may benefit from controlled growth environments to compensate for the narrowing of their defensive arsenals. (Summary by Mary Williams @PlantTeaching) Trends Plant Sci. 10.1016/j.tplants.2022.11.002
Plants synthesize interesting chemicals that attract, deter, amuse, and harm their predators. Some of the most harmful to humans have been selectively eliminated through the process of domestication, but others render potential food sources inedible. This review by Liu et al. discusses four approaches to increasing harvest index (product per plant) by selectively removing phytotoxins from otherwise edible tissues. As an example, antinutritional glucosinolates (GSLs) in canola seeds make the residue after extracting the oil inedible. Similarly, although cotton seeds have a lot of protein, they also contain phytotoxic gossypol, and cassava leaves contain cyanogenic glycosides. For these and other phytotoxins, several strategies have been explored to eliminate them from tissues that would otherwise be good food sources. These strategies include interfering with their biosynthetic enzymes or the genes that encode them, disrupting their transport or storage, or converting them to non-toxic forms. As the authors observe, detoxified plants may benefit from controlled growth environments to compensate for the narrowing of their defensive arsenals. (Summary by Mary Williams @PlantTeaching) Trends Plant Sci. 10.1016/j.tplants.2022.11.002
Review: How floral meristem termination shapes flowers
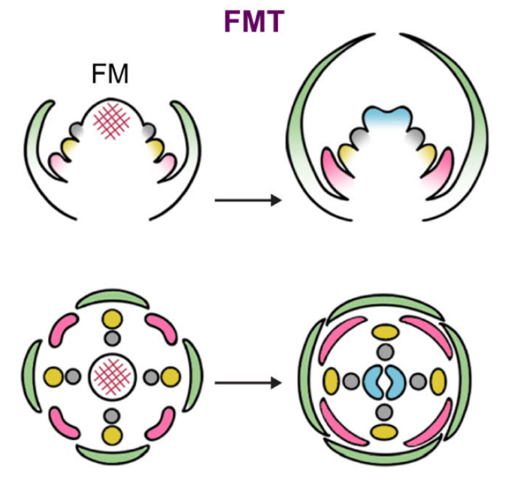 Flowers come in a breathtaking variety of shapes and sizes. The structures making up a flower, called sepals, petals, stamens, and carpels, all derive from the tightly regulated differentiation of a pool of stem cells in the so-called floral meristem. Floral meristem termination (FMT) is a crucial stage in floral development as it determines when stem cell proliferation in the flower meristem should stop. This is an essential and delicate step, and its regulation underlies the natural variation we observe in flowers. In this review, Min and Kramer first take an evolutionary perspective to highlight how variation in FMT can contribute to floral diversity through the control of whorl number (how many “layers” of floral organs make up a flower) and the number of floral organs per whorl (e.g. how many petals are there per “layer”). Then, the authors dive deeper into our current knowledge of the mechanisms regulating FMT at the molecular level, and they illustrate how changes in such mechanisms could result in a variety of different flower architectures. Finally, the authors advocate for a broader use of plant models with wide natural variation as a means to advance our understanding of FMT and unravel the molecular mechanisms that give rise to the countless different flowers that we humans love so much. (Summary by Laura Turchi @turchi_l) 10.1111/nph.18715
Flowers come in a breathtaking variety of shapes and sizes. The structures making up a flower, called sepals, petals, stamens, and carpels, all derive from the tightly regulated differentiation of a pool of stem cells in the so-called floral meristem. Floral meristem termination (FMT) is a crucial stage in floral development as it determines when stem cell proliferation in the flower meristem should stop. This is an essential and delicate step, and its regulation underlies the natural variation we observe in flowers. In this review, Min and Kramer first take an evolutionary perspective to highlight how variation in FMT can contribute to floral diversity through the control of whorl number (how many “layers” of floral organs make up a flower) and the number of floral organs per whorl (e.g. how many petals are there per “layer”). Then, the authors dive deeper into our current knowledge of the mechanisms regulating FMT at the molecular level, and they illustrate how changes in such mechanisms could result in a variety of different flower architectures. Finally, the authors advocate for a broader use of plant models with wide natural variation as a means to advance our understanding of FMT and unravel the molecular mechanisms that give rise to the countless different flowers that we humans love so much. (Summary by Laura Turchi @turchi_l) 10.1111/nph.18715
Roots of misinformation on common mycorrhizal networks in forests
 As an educator, I reply on metaphors and stories to engage my readers, yet the line between hyperbole and engagement is thin. The concept of plant-plant communication is exciting, and certainly has some truth, but unfortunately has spilled over into a narrative that perhaps is more fiction than fact. In particular, the notion that the soil mycorrhizal networks serve as conduits for communication and nutrient exchange has taken hold in the popular imagination. Here, Karst et al. systematically review evidence for this phenomenon, and find little evidence for it. They report that there is a citation bias towards reporting only the supporting data and omitting the caveats. Although informing the public of the value of forests and particularly old-growth forests is important, we as scientists must communicate with facts not fantasy. Furthermore, I’m not sure that the study of plants benefits when people try to ascribe them with human traits. As @NobodyNursery Tweeted, “I am in awe of the beauty of the cut-throat competition between plants on the forest floor, and I don’t need to anthropomorphize nature to respect and protect the ecology involved. We can love the forest even if the trees aren’t talking to each other.” Hear, hear! (Summary by Mary Williams @PlantTeaching). Nature Ecol. Evol. 10.1038/s41559-023-01986-1
As an educator, I reply on metaphors and stories to engage my readers, yet the line between hyperbole and engagement is thin. The concept of plant-plant communication is exciting, and certainly has some truth, but unfortunately has spilled over into a narrative that perhaps is more fiction than fact. In particular, the notion that the soil mycorrhizal networks serve as conduits for communication and nutrient exchange has taken hold in the popular imagination. Here, Karst et al. systematically review evidence for this phenomenon, and find little evidence for it. They report that there is a citation bias towards reporting only the supporting data and omitting the caveats. Although informing the public of the value of forests and particularly old-growth forests is important, we as scientists must communicate with facts not fantasy. Furthermore, I’m not sure that the study of plants benefits when people try to ascribe them with human traits. As @NobodyNursery Tweeted, “I am in awe of the beauty of the cut-throat competition between plants on the forest floor, and I don’t need to anthropomorphize nature to respect and protect the ecology involved. We can love the forest even if the trees aren’t talking to each other.” Hear, hear! (Summary by Mary Williams @PlantTeaching). Nature Ecol. Evol. 10.1038/s41559-023-01986-1
Anisotropic cell growth at the leaf base promotes age-related changes in leaf shape
 The Arabidopsis thaliana leaf exhibits dramatic phenotypic changes across the juvenile-to-adult phase transition during vegetative development. For example, juvenile leaves are small, round, and lack trichomes (leaf hairs). By contrast, adult leaves contain trichomes on the abaxial (or lower) leaf surface, serrations on the leaf margin, and a lengthened leaf blade. The switch between the juvenile and adult phases is regulated by the conserved miR156-SQUAMOSA PROMOTER BINDING PROTEINS-LIKE (SPLs) pathway. It has been shown that miR156 promotes the juvenile leaf phase. However, it is still unknown how SPLs specify adult leaf shape. Tang et al. used quantitative analyses of juvenile and adult leaves using MorphoLeaf software, overexpression lines, reporter lines, mutants, ChIP-seq, and computational modeling. They showed that miR156-targeted SPLs sufficiently promote cell expansion oriented along the abaxial leaf surface (cell growth anisotropy) and cell proliferation at the base of the blade which contributes to the adult leaf shape. In addition, ChIP-seq data shows that SPL10 might regulate leaf changes by activating genes involved in the cell cycle such as CYCLIND3 (CYCD3) and cell wall properties such as xyloglucan endotransglucosylase/hydrolase (XTH). Collectively, these results indicate that SPLs are important for the formation of the final shape of an adult leaf. (Summary by Andrea Gómez-Felipe @andreagomezfe) Plant Cell 10.1093/plcell/koad031
The Arabidopsis thaliana leaf exhibits dramatic phenotypic changes across the juvenile-to-adult phase transition during vegetative development. For example, juvenile leaves are small, round, and lack trichomes (leaf hairs). By contrast, adult leaves contain trichomes on the abaxial (or lower) leaf surface, serrations on the leaf margin, and a lengthened leaf blade. The switch between the juvenile and adult phases is regulated by the conserved miR156-SQUAMOSA PROMOTER BINDING PROTEINS-LIKE (SPLs) pathway. It has been shown that miR156 promotes the juvenile leaf phase. However, it is still unknown how SPLs specify adult leaf shape. Tang et al. used quantitative analyses of juvenile and adult leaves using MorphoLeaf software, overexpression lines, reporter lines, mutants, ChIP-seq, and computational modeling. They showed that miR156-targeted SPLs sufficiently promote cell expansion oriented along the abaxial leaf surface (cell growth anisotropy) and cell proliferation at the base of the blade which contributes to the adult leaf shape. In addition, ChIP-seq data shows that SPL10 might regulate leaf changes by activating genes involved in the cell cycle such as CYCLIND3 (CYCD3) and cell wall properties such as xyloglucan endotransglucosylase/hydrolase (XTH). Collectively, these results indicate that SPLs are important for the formation of the final shape of an adult leaf. (Summary by Andrea Gómez-Felipe @andreagomezfe) Plant Cell 10.1093/plcell/koad031
PIF4 regulates microtubule organization to mediate high temperature–induced hypocotyl elongation
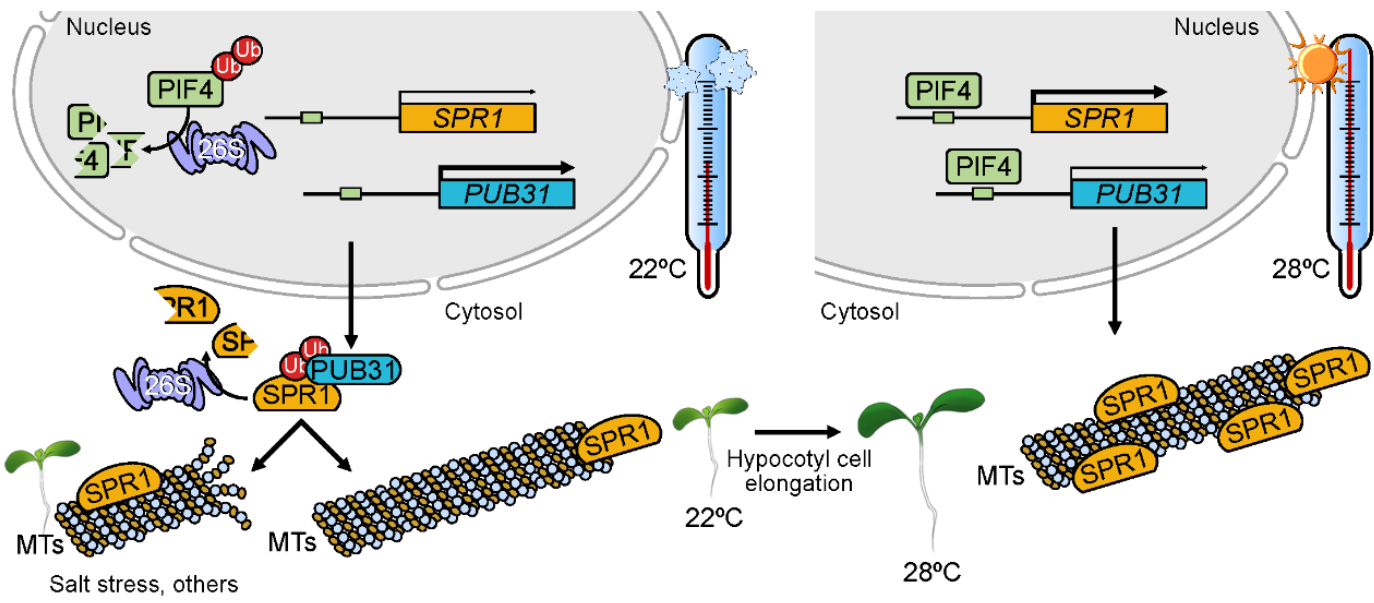 Plant growth adaptation to heat stress (thermomorphogenesis) is regulated by changes in plant morphology such as petiole and hypocotyl elongation. One of the known players in this response is PHYTOCHROME-INTERACTING FACTOR 4 (PIF4), a central regulator of hypocotyl elongation. However, the mechanisms underlying the response, including PIF4 interactors, are unclear. In addition, cortical microtubules can sense environmental signals and are related to the status of the of hypocotyl cells. In this work, Zhou et al. identified microtubule-associated proteins (MAPs), such as MAP SPIRAL1 (SPR1), involved in temperature-induced hypocotyl elongation. They used loss-of-function spr1 mutants of Arabidopsis thaliana, which showed shorter hypocotyls at 28 ºC. Expression of SPR1 increased with higher temperature in a PIF4-dependent manner. The authors not only proved the role of SPR1 but also highlighted the function of PLANT U-BOX TYPE E3 UBIQUITIN LIGASE 31 (PUB31), which ubiquitinates and degrades SPR1 when it is not necessary. In high temperature, PIF4 induces SPR1 which suppresses PUB31 expression, resulting in the accumulation of SPR1 that promotes hypocotyl cell elongation. This is one of the first works that fine tunes the mechanisms of microtubule modulation during heat stress in plants. (Summary by Eva Maria Gomez Alvarez, @eva_ga96) Plant Cell 10.1093/plcell/koad042
Plant growth adaptation to heat stress (thermomorphogenesis) is regulated by changes in plant morphology such as petiole and hypocotyl elongation. One of the known players in this response is PHYTOCHROME-INTERACTING FACTOR 4 (PIF4), a central regulator of hypocotyl elongation. However, the mechanisms underlying the response, including PIF4 interactors, are unclear. In addition, cortical microtubules can sense environmental signals and are related to the status of the of hypocotyl cells. In this work, Zhou et al. identified microtubule-associated proteins (MAPs), such as MAP SPIRAL1 (SPR1), involved in temperature-induced hypocotyl elongation. They used loss-of-function spr1 mutants of Arabidopsis thaliana, which showed shorter hypocotyls at 28 ºC. Expression of SPR1 increased with higher temperature in a PIF4-dependent manner. The authors not only proved the role of SPR1 but also highlighted the function of PLANT U-BOX TYPE E3 UBIQUITIN LIGASE 31 (PUB31), which ubiquitinates and degrades SPR1 when it is not necessary. In high temperature, PIF4 induces SPR1 which suppresses PUB31 expression, resulting in the accumulation of SPR1 that promotes hypocotyl cell elongation. This is one of the first works that fine tunes the mechanisms of microtubule modulation during heat stress in plants. (Summary by Eva Maria Gomez Alvarez, @eva_ga96) Plant Cell 10.1093/plcell/koad042
Mechanism behind ripened green bananas under high temperature
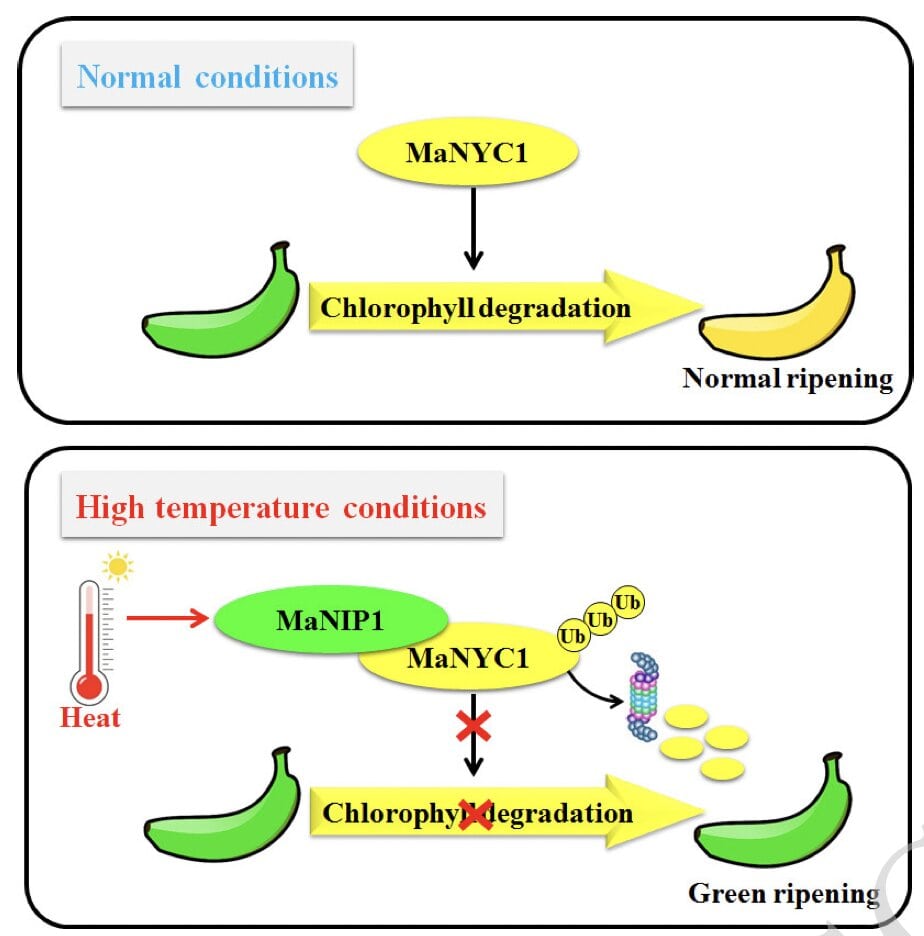 Heat stress is a common consequence of global warming for terrestrial plants, and it can have devastating effects on growth and yield. In terms of fruit ripening, high temperature is one of the most significant abiotic stresses inhibiting quality formation. During fruit ripening, chlorophyll breaks down in a normal process called “degreening,” which determines fruit quality and how nice the fruit looks. In most cases, high temperature stimulates degradation of chlorophyll. However, in the case of banana, which is the most popularly consumed fresh fruit worldwide, it can delay or completely abolish degreening. In this article, Luo and colleagues have deciphered the molecular mechanism behind the suppression of degreening at high temperatures in bananas. They identified a chlorophyll catabolic enzyme MaNYC1 (NON-YELLOW COLORING 1) that exhibited a reduced protein level when banana fruit were green ripening at high temperature. MaNYC1 is a well characterized NADPH-dependent chlorophyll b reductase in Arabidopsis. The authors found that NYC1 is rapidly degraded under high temperature and this degradation is facilitated by NYC1 interacting protein (NIP1) which essentially interacts with NYC1 and mediates its degradation via a proteasomal degradation pathway. Hence, this study provides an insight into a regulatory NYC1-NIP1 module in green ripening of bananas underlying fruit quality deterioration under high temperature stress. (Summary by Arpita Yadav @arpita_yadav_) 10.1093/plphys/kiad096
Heat stress is a common consequence of global warming for terrestrial plants, and it can have devastating effects on growth and yield. In terms of fruit ripening, high temperature is one of the most significant abiotic stresses inhibiting quality formation. During fruit ripening, chlorophyll breaks down in a normal process called “degreening,” which determines fruit quality and how nice the fruit looks. In most cases, high temperature stimulates degradation of chlorophyll. However, in the case of banana, which is the most popularly consumed fresh fruit worldwide, it can delay or completely abolish degreening. In this article, Luo and colleagues have deciphered the molecular mechanism behind the suppression of degreening at high temperatures in bananas. They identified a chlorophyll catabolic enzyme MaNYC1 (NON-YELLOW COLORING 1) that exhibited a reduced protein level when banana fruit were green ripening at high temperature. MaNYC1 is a well characterized NADPH-dependent chlorophyll b reductase in Arabidopsis. The authors found that NYC1 is rapidly degraded under high temperature and this degradation is facilitated by NYC1 interacting protein (NIP1) which essentially interacts with NYC1 and mediates its degradation via a proteasomal degradation pathway. Hence, this study provides an insight into a regulatory NYC1-NIP1 module in green ripening of bananas underlying fruit quality deterioration under high temperature stress. (Summary by Arpita Yadav @arpita_yadav_) 10.1093/plphys/kiad096
Inspired by nature: Self-burying seeds
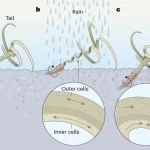
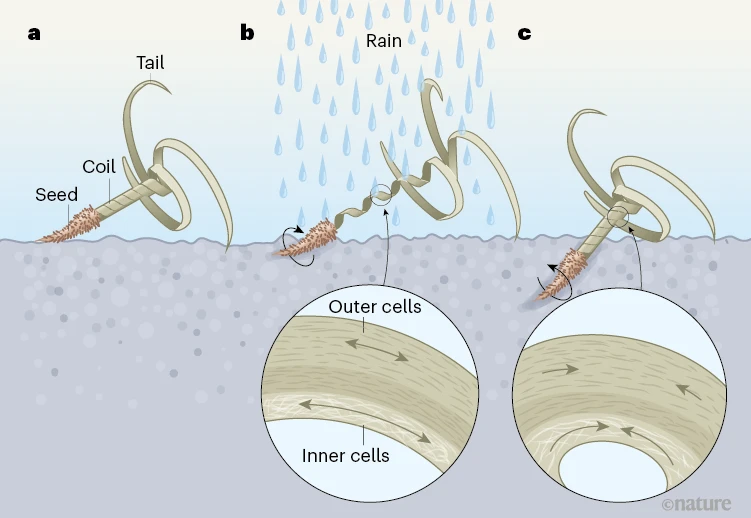 Self-burying seeds are high on the list of “aren’t plants amazing”. Seeds of several species carry appendages that change shape when exposed to moisture and that are oriented in such a way so that their shape changes push the seed underground (don’t take our word for it – see https://youtu.be/TOJG5mF6OLs). Burrowing underground gives the seed protection from being eaten by birds and more access to moisture, promoting survival and germination. Inspired by this, Luo et al. have developed an artificial carrier for seeds that confers to them this self-burying trait. The principle is hygromorphism – movement in response to water. Wood is hygromorphic as well as nontoxic and degradable, so this is a very clever strategy. The seeds can be sown onto the top of soil, and when their wooden carriers are wetted the “tails” extend, pushing the seed underground like the process that occurs with naturally self-burying seeds. The authors point out that these principles can also be used to push microorganisms and nutrients beneath the soil surface. Simple, effective, and elegant. (Summary by Mary Williams @PlantTeaching). Nature 10.1038/s41586-022-05656-3 See also the beautifully illustrated News and Views by Mason and Yakanama https://www.nature.com/articles/d41586-023-00396-4.
Self-burying seeds are high on the list of “aren’t plants amazing”. Seeds of several species carry appendages that change shape when exposed to moisture and that are oriented in such a way so that their shape changes push the seed underground (don’t take our word for it – see https://youtu.be/TOJG5mF6OLs). Burrowing underground gives the seed protection from being eaten by birds and more access to moisture, promoting survival and germination. Inspired by this, Luo et al. have developed an artificial carrier for seeds that confers to them this self-burying trait. The principle is hygromorphism – movement in response to water. Wood is hygromorphic as well as nontoxic and degradable, so this is a very clever strategy. The seeds can be sown onto the top of soil, and when their wooden carriers are wetted the “tails” extend, pushing the seed underground like the process that occurs with naturally self-burying seeds. The authors point out that these principles can also be used to push microorganisms and nutrients beneath the soil surface. Simple, effective, and elegant. (Summary by Mary Williams @PlantTeaching). Nature 10.1038/s41586-022-05656-3 See also the beautifully illustrated News and Views by Mason and Yakanama https://www.nature.com/articles/d41586-023-00396-4.